

Der Dünndarm (lateinisch Intestinum tenue) ist ein Teil des Verdauungstraktes und dient der Aufnahme von Nährstoffen aus der Nahrung. Dazu ist er mit zahlreichen Zotten (Erhebungen) und Krypten (Einsenkungen) ausgekleidet, so dass die Oberfläche stark vergrößert wird und ein Vielfaches der Körperoberfläche erreicht. Mit einer Länge von drei bis fünf Metern ist der Dünndarm der längste Teil des Verdauungstrakts. Er reicht vom Pförtner des Magens bis zur Ileozäkalklappe am Übergang zum Dickdarm und wird in den Zwölffingerdarm (Duodenum), den Leerdarm (Jejunum) und den Krummdarm (Ileum) gegliedert. Neben seiner Funktion zur Nährstoffaufnahme ist der Dünndarm auch der Hauptort der Wasserresorption des Menschen. Durchfallerkrankungen führen daher rasch zu Austrocknung.
Anatomie
Lage und Struktur

Lage und Struktur des Dünndarms unterscheiden sich je nach seinen Abschnitten: Er beginnt als Zwölffingerdarm (lat. Duodenum) mit einem erweiterten Abschnitt (Ampulla duodeni oder auch Bulbus duodeni) am Magenpförtner. Dieser ist ein Schließmuskel, der Musculus sphincter pylori, der die Aufgabe hat, den Speisefluss in den Zwölffingerdarm zu kontrollieren. Ist der Muskel in angespanntem Zustand, ist die Öffnung zwischen Magen und Zwölffingerdarm, das Ostium pyloricum, geschlossen und es kann kein Mageninhalt in den Zwölffingerdarm gelangen. Erst durch kurzzeites Entspannen und damit Öffnen des Ostium pyloricum kann Mageninhalt (Chymus) übertreten.
Der Zwölffingerdarm ist in etwa C-förmig und zwölf Fingerbreiten lang (daher der Name), was etwa 25 bis 30 cm entspricht. Der Chymus gelangt daher zuerst in einen oberen horizontalen Teil (Pars superior) in Höhe des ersten Lendenwirbelkörpers. Dieser Teil wird vom rechten Lappen der Leber überlagert, mit der er über ein Ligamentum hepatoduodenale verbunden ist. Außerdem berührt die Pars superior den Lobus quadratus der Leber und die Gallenblase.

Folgt man dem Chymus nun weiter im Zwölffingerdarm, kommt man in seinen absteigenden Anteil (Pars descendens), der bis zum dritten Lendenwirbelkörper hinabreicht. Dort münden beide Gänge der Bauchspeicheldrüse (Ductus pancreaticus und Ductus pancreaticus accessorius) und der Gallengang in den Zwölffingerdarm. Durch diese Gänge gelangen Gallenflüssigkeit und die Verdauungsenzyme der Bauchspeicheldrüse in den Darm, die nötig sind, um die Nahrung zu verdauen und aufzunehmen. Bei ihrer Mündung in den Darm werfen die Gänge Papillen auf, die Papilla duodeni minor und die Papilla duodeni major ("Vatersche Papille"). Neben der besonderen Nähe zur Bauchspeicheldrüse, tritt die Pars descendens in topographische Beziehung zur Niere und Nebenniere.
Dem Zwölffingerdarm weiter folgend, kommt man zu dessen zweitem horizontal verlaufenden Teil (Pars horizontalis) und einem letzten aufsteigenden Anteil (Pars ascendens). An einer Krümmung (der Flexura duodenojejunalis) tritt er schließlich in den Leerdarm (Jejunum) über. Dies markiert gleichzeitig das Ende des oberen Magen-Darm-Trakts. Hier bilden sich außerdem zwei Bauchfellnischen (Recessus duodenalis superior und inferior), in denen Teile des Dünndarms eingeklemmt werden können. Bei dieser, als Treitz-Hernie bezeichneten, Einklemmung kommt es zu einem lebensgefährlichen Darmverschluss.

Mit dem Jejunum beginnt der untere Magen-Darm-Trakt. Es geht ohne scharfe Grenze in den Krummdarm (Ileum) über. Beide liegen in Schlingen zwischen den Abschnitten des Dickdarms – Jejunum und Ileum werden sozusagen vom Dickdarm umrahmt. Zusammen machen sie mit 3 bis 5 Meter den größten Abschnitt des Dünndarms aus, wobei ca. 2/5 der Länge auf das Jejunum und 3/5 auf das Ileum entfallen. Ihre Länge schwankt, wie auch die Länge des gesamten Darms, von Mensch zu Mensch und hängt auch von der Spannung der Darmmuskulatur ab. In der Chirurgie rechnet man meist nur diese beiden Abschnitte zum Dünndarm. Das Ileum mündet schließlich End-zu-Seit mit dem Ostium ileale, auch genannt Ileozäkalklappe oder Bauhin-Klappe, in den Dickdarm.
Für die Lagebeziehungen des Dünndarms sind neben der Nachbarschaft zu anderen Organen vor allem die Peritonealverhältnisse entscheidend. So liegt der Zwölffingerdarm bis auf einen kleinen Abschnitt am Anfang sekundär retroperitoneal. Retroperitoneal heißt der Zwölffingerdarm ist nicht von Bauchfell (Peritoneum) umgeben; sekundär meint, dass dieser Zustand nicht von Anfang an so war, sondern dass es im Laufe der Entwicklung dazu kam. Jejunum und Ileum sind dagegen auf ganzer Länge intraperitoneal, also von Bauchfell umgeben. Eine besondere Bedeutung kommt dabei den Mesenterien zu: Dies sind Verdopplungen von Bauchfell, die von der hinteren Bauchwand zu den jeweiligen Organabschnitten ziehen. Im Bereich des Dünndarms entspringen die Mesenterien in Höhe der Lendenwirbelkörper drei bis fünf. Damit sind sie darmseitig deutlich länger als an ihrem Ursprung, weswegen die einzelnen Dünndarmschlingen gegeneinander verschieblich sind. In den Mesenterien befinden sich zudem die Leitungsbahnen (Blut-, Nerven- und Lymphgefäße) für Jejunum und Ileum.
Feinbau

Der Dünndarm weist grundsätzlich den typischen Wandbau des Magen-Darm-Trakts auf. Die innerste Schicht ist Schleimhaut (Mukosa) mit drei Unterabteilungen: Eine Schicht aus einschichtigem Zylinderepithel (Lamina epithelialis) stellt die innerste Barriere dar und liegt einer Schicht aus zellreichem Bindegewebe (Lamina propria) auf. Diese Schicht beherbergt zahlreiche Blutgefäße, Nerven, Lymphgefäße und Immunzellen. Die letzte Unterschicht der Schleimhaut setzt sich aus glatten Muskelzellen zusammen (Lamina muscularis mucosae), die der Schleimhaut Beweglichkeit verleihen. Der Schleimhaut schließt sich schließlich eine Schicht aus lockerem Bindegewebe (Tela submucosa) an, in dem sich die größeren Blut- und Lymphgefäße für die Schleimhaut und ein Nervengeflecht (Plexus submucosus) befinden. Außerdem ermöglicht sie der Schleimhaut Verschieblichkeit gegenüber der nachfolgenden Muskelschicht (Tunica muscularis). Diese besteht aus glatten Muskelzellen, die als innere Ringmuskel- (Stratum circulare) und als äußere Längsmuskelschicht (Stratum longitudinale) angeordnet sind. Diese Anordnung ermöglicht es den Organen des Magen-Darm-Trakts Pendel- und Segmentierungsbewegungen zur Durchmischung des Nahrungsbreis sowie peristaltische Bewegungen zum Weitertransport durchzuführen. Zwischen der Ring- und der Längsmuskelschicht befindet sich der Plexus myentericus (auch Auerbach-Plexus genannt), der ebenso wie der Plexus submucosus zum enterischen Nervensystem zählt. Die äußerste Schicht des Dünndarms ist das Bauchfell, das hier Serosa genannt wird. An den Abschnitten des Zwölffingerdarms, wo ein Überzug mit Bauchfell fehlt, gibt es stattdessen eine Schicht aus lockerem Bindegewebe, die Tunica adventitia.
Über diesen allgemeinen Aufbau des Magen-Darm-Trakts weist der Dünndarm einige wichtige Besonderheiten auf: Halbkreisförmige Falten, die Kerckring-Falten, oder Plicae circulares, sind quer zum Verlauf des Dünndarms angeordnet und springen in sein Inneres (Lumen) vor. Diese Falten werden von der Schleimhaut und der darunter liegenden Schicht, der Tela submucosa gebildet.
Von größerer Bedeutung ist das System aus Zotten und Krypten. Zotten sind fingerförmige Erhebungen der Dünndarmschleimhaut, die vor allem der Aufnahme von Bestandteilen der Nahrung dienen. Dazu besteht das Epithel der Zotten hauptsächlich aus Enterozyten – spezialisierte Zellen, die auf der zum Lumen gerichteten Seite zahlreiche Mikrovilli und spezielle Transporter tragen. Die Mikrovilli vergrößern die Darmoberfläche extrem zur besseren Resorption; die Transporter transportieren direkt Nahrungsbestandteile in die Zelle. Zum Abtransport der aufgenommenen Stoffe (außer Fette) durchziehen die Zotte in Längsrichtung (also nach innen, zum Lumen hin) mehrere Arteriolen. An der Spitze gehen sie in ein flächiges System aus Kapillaren unter dem Epithel über. Durch dieses Kapillarsystem können die Stoffe abtransportiert werden. Dazu gelangen sie zunächst in die Kapillaren, die dann in eine zentrale Venole gelangen. Die Venole leitet das Blut und die Stoffe aus der Nahrung weiter in Gefäßsysteme unter der Schleimhaut und weiter über die Venen des Dünndarms zur Pfortader (siehe auch Abschnitt Blutversorgung und Lymphabfluss). Die Fette aus der Nahrung werden hingegen über die Lymphe abtransportiert, so dass in den Zotten auch eine oder mehrere Lymphekapillaren verlaufen. Diese transportieren die Lymphe zu größeren Lymphgefäßen an der Darmwand.
Krypten sind im Gegensatz zu Zotten Einsenkungen des Epithels in der Schleimhaut. An der Basis dieser Krypten sitzen multipotente Stammzellen, die sich fortlaufend teilen. Ihre Abkömmlinge steigen an der Kryptenwand bis zur Spitze der Zotten empor, differenzieren sich während ihrer Wanderung und nehmen ihre spezifischen Aufgaben wahr. An der Zottenspitze schließlich sterben die Zellen entweder durch Zelltod oder werden abgestoßen. So werden die Epithelzellen der Zotten etwa alle fünf Tage komplett ausgetauscht, wobei die Zellerneuerung von den Krypten her beginnt. Außerdem beinhalten die Krypten Paneth-Zellen. Diese Zellen sind für die Immunabwehr verantwortlich, wofür sie antibakterielle Eiweiße ausscheiden. Die Zotten und Krypten vergrößern die Oberfläche des Dünndarms auf das etwa 7 bis 14-fache (ca. 4 m²). Rechnet man den Bürstensaum (Gesamtheit der Mikrovilli) hinzu, schätzt man die Darmoberfläche auf ca. 60 bis 200 m² (je nach Lehrbuch), die damit wesentlich größer ist als die Körperoberfläche.
Eine weitere Besonderheit des Dünndarms, die aber nur im Zwölffingerdarm zu finden ist, sind die Brunner-Drüsen (Glandulae duodenales). Diese Drüsen liegen in großen Paketen (die im Verlauf des Dünndarms immer kleiner werden) in der Submukosa unter der Schleimhaut. Sie sind am Aufbau der Schleimschicht beteiligt, die das Epithel gegen den sauren Mageninhalt schützt und scheiden Hydrogencarbonat aus, das der Neutralisation des Mageninhalts dient.
Blutversorgung und Lymphabfluss

Im Verlauf des Magen-Darm-Trakts ändert sich die Versorgung der einzelnen Organe. Dem Zwölffingerdarm als Übergangsstruktur zwischen oberem und unterm Magen-Darm-Trakt kommt daher eine besondere Rolle zu: Äste aus dem Bauchhöhlenstamm (Truncus coeliacus) der Aorta gehen mit Ästen aus der oberen Eingeweidearterie (Arteria mesenterica superior) eine Verbindung (Anastomose) ein. Der Truncus coeliacus entsendet eine gemeinsame Leberarterie (Arteria hepatica communis), der eine Arterie für Magen und Zwölffingerdarm entspringt (Arteria gastroduodenalis). Aus der Arteria gastroduodenalis wiederum gehen die obere Bauchspeicheldrüsen-Zwölffingerdarm-Arterie (Arteria pancreaticoduodenalis superior) und die hintere Bauchspeicheldrüsen-Zwölffingerdarm-Arterie (Arteria pancreaticoduodenalis posterior) ab, die mit direkten Ästen (Rami duodenales) den oberen und den absteigenden Teil des Zwölffingerdarms versorgen. Die obere Bauchspeicheldrüsen-Zwölffingerdarm-Arterie verbindet sich (anastomosiert) mit der unteren Bauchspeicheldrüsen-Zwölffingerdarm-Arterie, die aus der oberen Eingeweide-Arterie (Arteria mesenterica superior) stammt, und die ihrerseits mit Ästen die weiteren Abschnitte des Zwölffingerdarms vorsorgt. Die Venen verlaufen wie die Arterien und sind wie diese benannt, das heißt, es gibt auch hier eine untere und eine obere Bauchspeicheldrüsen-Zwölffingerdarm-Vene, die miteinander anastomosieren. Sie münden entweder direkt in die Pfortader (Vena portis hepatis) oder über die obere Eingeweidevene (Vena mesenterica superior). Varianten sind jedoch häufig.

Leer- und Krummdarm werden dagegen nur durch Äste der oberen Eingeweide-Arterie (Arteria mesenterica superior) versorgt. Leerdarm- und Krummdarm-Arterien (Arteriae jejunales und Arteriae ileales) gehen aus ihr ab und bilden drei übereinander liegende Gefäßarkaden (bogenförmig verlaufende Gefäßabschnitte) aus. Von den Arkaden gehen schließlich die Endarterien, die Arteriae rectae ab, die mit geradem Verlauf zur Darmwand ziehen. Die Venen sind gleich wie die Arterien angeordnet und münden alle in die obere Eingeweidevene (Vena mesenterica superior), die in die Pfortader der Leber mündet.
Die Lymphe des Dünndarms hat die besondere Aufgabe, Fette (Lipide), die von den Darmzotten aufgenommen wurden, abzutransportieren. Der Lymphabfluss von Zwölffingerdarm und Jejunum und Ileum ist dabei unterschiedlich. Die Lymphe des Zwölffingerdarms gelangt zunächst in die oberen und unteren Bauchspeicheldrüsen-Zwölffingerdarms-Lymphknoten (Nodi lymphoidei pancreaticoduodenales superiores und inferiores) und von dort weiter in Lymphknoten rund um den Bauchhöhlenstamm, die Nodi lymphoidei coeliaci. Anschließend fließt sie weiter ab in den Darmstamm (Truncus intestinalis), in die Lendenzisterne (Cisterna chyli) und schließlich in den Milchbrustgang (Ductus thoracicus), der schlussendlich im linken Venenwinkel endet. Die zahlreichen Lymphkapillaren aus den Zotten von Jejunum und Ileum (siehe Abschnitt Feinbau) vereinigen sich zu größeren Lymphkapillaren. Diese ziehen mit den Blutgefäßen in den Gefäßarkaden und gelangen zu 100 bis 200 Lymphknoten, den Nodi lymphoidei juxtaintestinales. Von dort fließt die Lymphe in die oberen Eingeweide-Lymphknoten (Nodi mesenterici superiores), weiter in den Darmstamm und dann weiter wie die Lymphe des Zwölffingerdarms bis zum linken Venenwinkel. Von dort aus gelangt sie über die rechte Herzhälfte zuerst in die Lunge, was sich dort zur Synthese der Surfactants als sinnvoll erweist.
Innervation
Der gesamte Dünndarm wird parasympathisch vom hinteren Stamm des Vagusnervs (Truncus vagalis posterior) innerviert. Sympathische Nervenfasern für den Zwölffingerdarm stammen aus dem großen Eingeweidenerv (Nervus splanchnicus major) aus den Rückenmarkssegmenten Th5 bis Th9 und laufen zur Aorta, wo sie zusammen mit anderen Nervenfasern ein Nervengeflecht, den Plexus aorticus abdominalis bilden. In dieses Nervengeflecht eingelagert liegt das Ganglion coeliacum, wo die Nervenfasern für das Duodenum umgeschaltet werden und von dort weiter zum Zwölffingerdarm ziehen. Leer- und Krummdarm werden durch sympathische Fasern versorgt, die als kleiner Eingeweidenerv (Nervus splanchnicus minor) aus den Rückenmarkssegmenten Th10 und 11 laufen, ebenfalls zum Plexus aorticus abdominalis ziehen und dort im Ganglion mesentericum superius umgeschaltet werden. Von dort erreichen sie ihre Erfolgsorgane, Jejunum und Ileum.
Entwicklung und Entwicklungsstörungen
Die einzelnen Anteile des Dünndarms entwickeln sich im Embryo aus unterschiedlichen Teilen des primitiven Darmrohrs: Der obere horizontale Teil des Zwölffingerdarms, die Pars superior entwickelt sich noch aus dem Vorderdarm, während alle übrigen Teile des Zwölffingerdarm wie auch Jejunum und Ileum aus dem Mitteldarm stammen. Anfangs verfügen noch alle Teile des Darms über einen Überzug mit Bauchfell, also über ein Mesenterium, das von der Rückwand der Bauchhöhle an die jeweiligen Organe herantritt. Zusätzlich verfügt die Pars superior des Zwölffingerdarms, wie der Magen auch, noch über ein weiteres Mesenterium, das von der Vorderseite der Bauchhöhle kommt. Die Pars superior des Zwölffingerdarms wird also von einem vorderen und einem hinteren Mesenterium, die übrigen Abschnitte des Darms nur von einem hinteren Mesenterium bedeckt. Die Mesenterien der jeweiligen Organe in der Entwicklung heißen wie folgt:
| vorderes (ventrales) Mesenterium | Organ(abschnitt) | hinteres (dorsales) Mesenterium |
|---|---|---|
| Mesogastricum ventrale | Magen | Mesogastricum dorsale |
| Mesoduodenum ventrale | Zwölffingerdarm, Pars superior | Mesoduodenum dorsale |
| – | Zwölffingerdarm, Pars descendens, horizontalis und ascendens | |
| – | Jejunum und Ileum | Mesenterium |
| – | Blinddarm | Mesocaecum |
Außerdem entwickeln sich aus dem Epithel des Zwölffingerdarms verschiedene Organe in die Mesenterien hinein: Leber und Gallenwege entwickeln sich in das Mesogastricum ventrale und Mesoduodenum ventrale, vordere Bauchspeicheldrüsenanlage ins Mesoduodenum ventrale und hintere Bauchspeicheldrüsenanlage ins Mesoduodenum dorsale.
Für die weitere Entwicklung des Zwölffingerdarms ist die Drehung des Magens von entscheidender Bedeutung, die etwa ab der fünften Entwicklungswoche stattfindet. Dabei dreht sich der Magen von oben betrachtet im Uhrzeigersinn um etwa 90° und kippt schließlich um eine sagittale Achse (Achse von vorn nach hinten). Dadurch gelangt der Zwölffingerdarm erst nach rechts und dann etwas nach oben und entwickelt bis zum Abschluss der Magendrehung seine typische C-Form. Der Zwölffingerdarm dreht sich außerdem nach hinten zur Bauchwand, und sein Überzug mit Bauchfell (Peritoneum viscerale) verschmilzt mit dem Bauchfell der hinteren Bauchhöhle (Peritoneum parietale), so dass er (bis auf einen kleinen Teil der Pars superior) nicht mehr von Bauchfell überzogen ist, er wird sekundär retroperitonealisiert.
Gleichzeitig mit der Drehung des Magens im Bereich des Vorderdarms findet im Bereich des Mittel- und Hinterdarms eine andere Drehung statt: Die der Nabelschleife. Dabei dreht sich das gesamte, schleifenförmige Darmrohr zwischen der sechsten und elften Entwicklungswoche in diesem Bereich um eine Achse, die von der oberen Eingeweidearterie (Arteria mesenterica superior) und dem Dottergang (Ductus omphaloentericus) gebildet wird. Von vorne betrachtet dreht sich das Darmrohr um insgesamt 270° gegen den Uhrzeigersinn. Damit gelangt der ehemals obere Teil der Nabelschleife, der mundzugewandte (orale) Teil, nach unten, und der ehemals untere Teil, mundabgewandte (aborale) Teil nach oben. Da sich aus mundzugewandtem Teil Jejunum und Ileum bilden, und aus dem mundabgewandten Teil der Blind- und Dickdarm entsteht, führt dies zur typischen Lage von Jejunum und Ileum zum Dickdarm (der Dickdarm bildet einen Rahmen um Ileum und Jejunum). Das Darmrohr wächst während dieser Drehung stark in die Länge und zwar der mundwärtige Teil der Nabelschleife deutlich stärker als der mundabgewandte. Wegen des unterschiedlichen Längenwachstums des mundwärtigen und mundabgewandten Teils der Nabelschleife ist der Dünndarm wesentlich länger als der Dickdarm. Daraus resultiert ein Platzmangel, so dass sich Jejunum und Ileum in zahlreiche Schlingen zwischen dem Rahmen des Dickdarms legen muss. Der erste Abschnitt der Nabeldrehung (die ersten 90°) findet im Dottersack, also außerhalb des Embryos, statt (physiologischer Nabelbruch). Diese ausgelagerten Darmschlingen werden erst in der zehnten Entwicklungswoche in den Embryo zurückverlagert. Findet diese Rückverlagerung nicht statt, kommt es zur Omphalozele.
Eine weitere Entwicklungsstörung ist eine Malrotation, bei der die Drehung der Nabelschleife nicht oder unvollständig abläuft mit der Folge, dass Darmteile in völlig untypischer Lage zum Liegen kommen können. Eine Malrotation kann völlig unbemerkt bleiben, sie kann aber auch zu einem Volvulus oder anderen Formen der Strangulation von Darmabschnitten (Duodenalstenose, Arteria-mesenterica-superior-Syndrom) führen. Daneben kann das Dünndarmlumen nicht durchgängig sein (Dünndarmatresie), wenn es zum Beispiel durch eine Membran verschlossen oder nicht auf ganzer Länge ausgebildet ist. Zudem kann der Dottergang nicht richtig veröden und ein Meckel-Divertikel bilden.
Funktion
Da der Speisebrei (Chymus) aus dem Magen ausgesprochen sauer ist (der pH-Wert im Magen liegt unter 2), muss dieser erst neutralisiert werden. Das geschieht im Wesentlichen im Zwölffingerdarm, indem ein neutralisierendes Sekret abgegeben wird (Sekretion).
Der gesamte Dünndarm ist weiterhin Hauptort der Verdauung und der Aufnahme der Nahrungsbestandteile (Kohlenhydrate, Eiweiße, Fette, Vitamine, Elektrolyte und Wasser), für das Ileum kommt noch die Aufgabe der Immunabwehr (durch die Peyer-Plaques) hinzu.
Sekretion
Die Neutralisation des sauren Chymus erfolgt über Hydrogencarbonat (Bikarbonat), das im Dünndarm von den Brunner-Drüsen des Zwölffingerdarms, von den Epithelzellen der Krypten und von Enterozyten abgegeben wird. Daneben gelangt alkalisches, Hydrogencarbonat-reiches Sekret aus der Bauchspeicheldrüse über die beiden Ausführungsgänge der Bauchspeicheldrüse (Ductus pancreaticus und Ductus pancreaticus accessorius) in den Dünndarm und trägt so zur Neutralisation der Säure bei. Durch die Neutralisation können die Verdauungsenzyme optimal arbeiten. Die Steuerung der Sekretion erfolgt über lokale Reflexe, die über das enterische Nervensystem vermittelt werden und über verschiedene Hormone wie CCK, Sekretin, Gastrin.
Daneben sezernieren die Epithelzellen der Krypten eine Natriumchlorid-haltige Flüssigkeit und die Becherzellen Muzine, die die Schleimhaut des Dünndarms als Unstirred Layer überziehen. Dieser dient dem Schutz der Schleimhaut und als Gleitschicht, damit der Nahrungsbrei vorbeigleiten kann.
Verdauung
Zur Verdauung zählt man prinzipiell die Aufspaltung der Nahrung in ihre verschiedenen Bestandteile (genauer: den Abbau hin zu niedermolekularen Verbindungen) und deren Aufnahme. Dazu dienen die Bewegungen der Organe des Magen-Darm-Trakts, die man als gastrointestinale Motilität bezeichnet, weiterhin die verschiedenen Verdauungssekrete aus den Mundspeicheldrüsen, dem Magen, dem Pankreas und der Leber und schließlich die Verdauungsenzyme, die sich in den Verdauungssekreten und im Bürstensaum des Darms befinden.
Motilität des Dünndarms
Die Bewegungen (Motilität) des Dünndarms transportieren die Nahrung im Dünndarm weiter, zerkleinern sie und vermischen sie mit den Verdauungssekreten. Im Folgenden ist zunächst die Motilität des Dünndarms während der Verdauungsphase und zwischen den Verdauungsphasen beschrieben, die neuronale Grundlage, die wesentlich komplexer ist, folgt danach.
In der Verdauungsphase (digestive Phase) kommt es zu Segmentations- und Pendelbewegungen des Dünndarms. Darunter versteht man Kontraktionen des Dünndarms, die in erster Linie der Durchmischung des Nahrungsbreis und dem Kontakt des Nahrungsbreis mit der Darmwand dienen und nicht so sehr dem Transport der Nahrung. Genauer gesagt kontrahiert sich bei Segmentationsbewegungen die Ringmuskulatur in benachbarten Bereichen des Dünndarms, so dass quasi einzelne Segmente erkennbar sind. Bei Pendelbewegungen kommt es zu einer Längsverschiebung der Darmwand über den Darminhalt hinweg, ausgelöst durch Kontraktionen der Längsmuskulatur. Auslöser dieser Bewegungen sind Schrittmacherzellen, die Cajal-Zellen, und die Frequenz der Schrittmacherzellen beträgt im Zwölffingerdarm noch 12 pro Minute und nimmt zum Ileum auf 8 pro Minute hin ab.
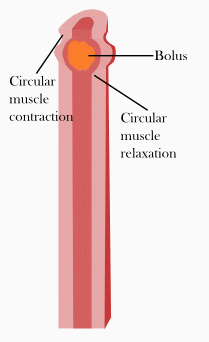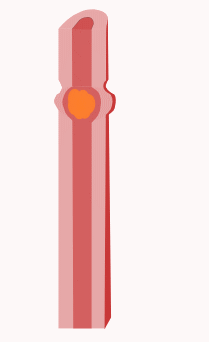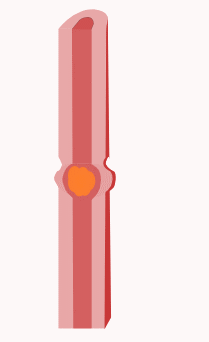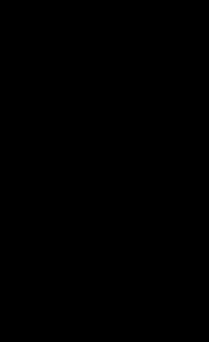
Die Segmentations- und Pendelbewegungen werden lokal von einer propulsiven Peristaltik überlagert. Propulsive Peristaltik meint Kontraktionen der Muskulatur, die dazu führen, dass der Nahrungsbrei weitertransportiert wird, also nach aboral (mundfern). Dazu kontrahiert sich die Ringmuskulatur oral (mundnah) vom Nahrungsbrei und drückt diesen somit nach aboral. Gleichzeitig entspannt sich die Ringmuskulatur aboral vom Nahrungsbrei und die Längsmuskulatur aboral kontrahiert sich. Das führt dazu, dass sich das Darmlumen aboral erweitert, dass dem Nahrungsbrei also aboral Platz geschaffen wird und dieser somit weiter transportiert werden kann. Je nach Nahrungszusammensetzung dauert der Transport bis zum Blinddarm etwa zwei bis zehn Stunden.
Zwischen den Verdauungsphasen (interdigestive Phase) treten mehrere einzelne Phasen auf: Nach einer Ruhezeit von ca. einer Stunde (Phase 1) kommt es etwa 30 min lang zu ungerichteten Kontraktionen des Dünndarms (Phase 2). Im Anschluss wandern ca. 15 min lang motorische Wellen, MMC (Migrating Motor Complex), über den Dünndarm. Das sind kräftige peristaltische Wellen, also Kontraktionen, die im Magen oder im Zwölffingerdarm beginnen und vor dem Dickdarm enden. Ziel dieser motorischen Wellen ist es, unverdaute Reste aus dem Dünndarm zu entfernen und überschießendes Bakterienwachstum zu verhindern. Auch sie werden von den Cajal-Schrittmacherzellen ausgelöst und ihre Frequenz beträgt etwa drei pro Minute. Nach den motorischen Wellen folgt eine vierte Phase, in der die Aktivität wieder abnimmt, bis es schließlich wieder zur Ruhephase (Phase 1) kommt. Der Übergang von der zweiten zur dritten Phase scheint vom Hormon Motilin mitgesteuert zu werden, der Übergang von der interdigestiven zur digestiven Phase wird dagegen vom Vagusnerv gesteuert.
Am Übergang vom Dünndarm zum Dickdarm befindet sich die Ileozäkalklappe, durch die täglich etwa zwei Liter flüssiger Dünndarminhalt hindurchtreten. Die Klappe ist prinzipiell verschlossen und öffnet sich erst bei einem Druckanstieg im Ileum, dem Dünndarmabschnitt vor der Klappe. Es tritt dann folglich Dünndarminhalt in den Dickdarm über. Bei einem Druckabfall im Ileum verschließt sie sich hingegen, so dass der Übertritt von Dickdarminhalt in den Dünndarm verhindert wird und damit der bakterienreiche Dickdarm vom bakterienärmeren Dünndarm getrennt ist.
Es folgen nun die neuronalen Grundlagen für die Dünndarmmotilität:
Ab dem Magen findet man in allen Organen einen langsamen basalen elektrischen Rhythmus, das heißt, man findet langsame Potentialschwankungen der Muskulatur mit niedriger Frequenz, sogenannte slow waves. Im Dünndarm beträgt die Frequenz dieser slow waves ca. 12 pro Minute. Auslöser für diese Potentialschwankungen sind Schrittmacherzellen, die Cajal-Zellen. Sie haben Verbindungen zu den benachbarten Muskelzellen (Gap junctions), über die die Potentialschwankungen weitergeleitet werden, und bilden so ein Netzwerk zwischen der Ring- und der Längsmuskulatur des Dünndarms (für die Beschreibung der Dünndarmmuskulatur siehe Dünndarm#Feinbau). Kommt es nun zur Dehnung der Darmwand oder werden bestimmte Hormone oder Neurotransmitter ausgeschüttet, kommt es zu wesentlich schnelleren Potentialschwankungen mit größerer Amplitude, den Spike-Potentialen, im betreffenden Abschnitt des Dünndarms. Je nach Frequenz dieser Spike-Potentiale kontrahiert sich die Wandmuskulatur unterschiedlich stark.
Daneben wird die Motilität des Dünndarms von Nerven und Hormonen gesteuert. Es sind viele verschiedene Hormone beteiligt. Sie werden zu bestimmten Zeiten bei der Verdauung freigesetzt und regulieren den Gesamtablauf, z. B. wann welche Verdauungssäfte freigesetzt werden. Der Dünndarm verfügt – wie auch andere Teile des Verdauungssystems – über ein eigenes Nervensystem, das enterische Nervensystem. Es umfasst zwei Nervengeflechte: den Auerbach-Plexus und den Meissner-Plexus, die miteinander verbunden sind. Der Auerbach-Plexus reguliert dabei vor allem die Durchblutung und die Motilität, während der Meissner-Plexus eher die Sekretion steuert. Die Motilität kann vom enterischen Nervensystem völlig autonom geregelt werden. Wird zum Beispiel ein Darmabschnitt durch den Darminhalt gedehnt, so wird diese Dehnung von Sensoren des Meissner-Plexus registriert und über weitere Nervenzellen wird diese Information an den Auerbach-Plexus weitergegeben, der bewirkt, dass sich die Darmmuskulatur mundwärts (oral) vom Speisebrei anspannt, während sich die vom Speisebrei mundferne (aborale) Muskulatur entspannt. Dadurch wird die Nahrung quasi nach aboral gedrückt, also weitertransportiert (propulsive Peristaltik). Dies bezeichnet man auch als peristaltischen Reflex.
In die Regulation durch das enterische Nervensystem kann durch den Sympathikus und den Parasympathikus eingegriffen werden (extrinsische Innervierung). Dabei hemmt der Sympathikus die Darmmotilität, wohingegen der Parasympathikus sie fördert. Die sympathischen Fasern stammen aus den Brust- und Lendenabschnitten des Rückenmarks und werden in Ganglien (Nervenzellanhäufungen) nahe dem Rückenmark auf weitere Nervenfasern (sogenannte postganglionäre Fasern) umgeschaltet, die zu den Zielzellen im Darm ziehen. Der Transmitter für die Übertragung auf die Zielzellen ist Noradrenalin. Die Zielzellen sind überwiegend erregende Neurone des enterischen Nervensystems, teilweise aber auch direkt Muskelzellen. Die Rezeptoren der Muskulatur sind bei der longitudinalen Muskulatur α2-Adrenozeptoren (mit Gi als signaltransduzierendes G-Protein) und bei der zirkulären Muskulatur α1-Adrenozeptoren (mit Gq als signaltransduzierendes G-Protein). Die parasympathischen Fasern stammen hingegen aus der Medulla oblongata und ziehen mit dem Vagusnerv zum Dünndarm.
Von einigen Nervenfasern liegen Endigungen frei in der Darmwand. Diese Nerven fungieren als Sensoren für mechanische, chemische und Schmerzreize und ziehen mit den Fasern von Sympathikus und Parasympathikus zum zentralen Nervensystem. So können vagovagale Reflexe entstehen.
Enzymatische Verdauung
Im Dünndarm wird die Nahrung enzymatisch verdaut. Dazu dienen Verdauungsenzyme im Mund (z. B. Amylasen), Magen (die Peptidasen) und später auch der Bauchspeicheldrüse (z. B. Pankreaslipase). Durch die Verdauungsenzyme werden die Kohlenhydrate, Proteine und Fette in ihre Bestandteile aufgespalten, Proteine jedoch bevorzugt nicht in einzelne Aminosäuren, sondern in Di- und Tripeptide (Moleküle aus zwei oder drei Aminosäuren). Im Dünndarm wirken die Maltase-Glucoamylase, Laktase und Saccharase-Isomaltase, die die Spaltung von α-1,4-glykosidischen Bindungen der Kohlenhydratketten bewirkt. Das heißt, die Kohlenhydrate werden in ihre Einfachzucker (Monosaccharide) aufgespalten. Diese Enzyme sitzen in den Membranen der Dünndarmzellen des Bürstensaums, man zählt sie daher zur membranassoziierten Verdauung.
Die Pankreasenzyme gelangen durch den Ductus pancreaticus über die Papilla duodeni major in das Duodenum. Die Galle dient der Emulgierung der Fette (siehe Aufnahme der Fette). Zusätzlich wird der Galle das Bilirubin und andere Stoffe, die ausgeschieden werden sollen, beigemischt und damit ausgeschieden. Die Gallensäure hingegen wird zu rund 95 % resorbiert und wieder der Leber zur Verfügung gestellt.
Aufnahme der Nahrung im Dünndarm
Der Dünndarm ist hauptsächlich für die Absorption von Kohlenhydraten, Proteinen und Fetten verantwortlich – Wasser und Elektrolyte werden auch im Dickdarm absorbiert. Die Aufnahme von Nahrungsstoffen wird durch die große Oberfläche des Dünndarms begünstigt und geschieht im Wesentlichen über Transportvorgänge in die Enterozyten. Dazu erzeugen die Enterozyten einen Konzentrationsunterschied, einen Gradienten von Natrium und Kalium gegenüber ihrer Umgebung. Dieser Gradient ist die treibende Kraft, um Nahrungsbestandteile in die Dünndarmzelle zu transportieren. Man unterscheidet zwei Formen der Stoffaufnahme: den transzellulären Transport und den parazellulären Transport. Unter transzellulärem Transport versteht man die Aufnahme über spezielle Transportproteine. Das heißt, diese speziellen Transportproteine transportieren die Stoffe in die Zelle hinein, unter Umständen auch entgegen ihrem Gradienten (Konzentrationsgefälle). Der parazelluläre Transport hingegen erfolgt entlang eines Gradienten zwischen den Zellen hindurch.
Aufnahme von Kohlenhydraten

Kohlenhydraten können nur als Einfachzucker (Monosaccharide) aufgenommen werden. Der Großteil wird im Zwölffingerdarm und Leerdarm absorbiert – ein kleiner Teil erreicht den Dickdarm und wird von dort ansässigen Bakterien verstoffwechselt. Es gibt zwei Transportproteine für die Absorption der Monosaccharide:
- Glukose und Galaktose werden zusammen mit Natrium (Natrium-Symport) über den Transporter SGLT1 (Sodium Glucose Transporter 1) in den Enterozyten aufgenommen. Natrium folgt dabei seinem Gradienten in die Zellen hinein und treibt den Transport von Glukose/Galaktose an.
- Fruktose gelangt über erleichterte Diffusion durch den Transporter GLUT5 (Glucose Transporter 5) in die Zelle, das heißt, der Transporter erleichtert den „Fluss“ von Fructose, übernimmt aber keinen aktiven Transport.
Alle drei Zucker gelangen dann durch den Transporter GLUT2 (Glucose Transporter 2) ins Blut. Triebkraft für den Transport ins Blut ist der Gradient: Im Enterozyten ist die Konzentration der drei Monosaccharide also vergleichsweise hoch und im Blut vergleichsweise gering, so dass die Monosaccharide einfach in Blut diffundieren können.
Aufnahme von Proteinen

Auch Proteine müssen prinzipiell zuerst in ihre Aminosäuren oder in Di- oder Tripeptide gespalten werden, bevor sie aufgenommen werden. Während Säuglinge Proteine noch in nennenswerter Anzahl unverdaut über Endozytose aufnehmen können, ist dieser Mechanismus beim Erwachsenen praktisch unbedeutend. Insgesamt werden im Zwölffingerdarm und im Leerdarm etwa 80 bis 90 % der Proteine resorbiert; nur 10 % gelangen in den Dickdarm, wo sie von Bakterien verstoffwechselt werden. Somit werden letztlich über 96 % des in der Nahrung enthaltenen Stickstoffs (überwiegend in Proteinen) aufgenommen.
- Einzelne Aminosäuren gelangen über verschiedene Natrium-Symporter in den Enterozyten und verlassen diesen wieder über erleichterte Diffusion und gelangen ins Blut.
- Di- und Tripeptide werden durch den Transporter PepT1 zusammen mit einem Proton in die Zelle aufgenommen. In der Zelle werden die Di- und Tripeptide überwiegend dann in einzelne Aminosäuren aufgespalten und verlassen die Zelle über erleichterte Diffusion ins Blut. Einige wenige Di- und Tripeptide können auch ungespalten ins Blut gelangen.
- Der für den PepT1-Transporter notwendige Gradient an Protonen wird vom Transporter NHE3 aufgebaut, der Protonen aus der Zelle ins Dünndarmlumen und Natriumionen aus dem Dünndarmlumen in die Zelle transportiert. Da der dafür nötige Natrium-Gradient wiederum von der Natrium-Kalium-ATPase aufgebaut wird, handelt es sich beim Transport von Di- und Tripeptiden um einen tertiär aktiven Transport.
Aufnahme von Fetten
Der Großteil (ca. 95 %) der Nahrungsfette wird im Zwölffingerdarm und Leerdarm aufgenommen, einige kurzkettige Fettsäuren darüber hinaus auch im Kolon. Nur etwa fünf bis sieben Gramm Fett werden mit dem Stuhl bei durchschnittlicher Ernährung ausgeschieden. Enzyme des Speichels (Zungengrundlipase), des Magens (Magenlipase) und der Bauchspeicheldrüse (Pankreaslipase) spalten die Fette, Fettester und Cholesterinester in Cholesterin, Monoglyceride, Fettsäuren und Lysophospholipide (Lipolyse), die vom Dünndarm aufgenommen werden können. Dabei bilden die genannten Spaltprodukte mit den Gallensäuren aus der Galle sogenannte gemischte Mizellen. In den Mizellen kommen darüber hinaus Triacylglyceride und fettlösliche Vitamine und Phospholipide vor. Effektiv resorbiert werden alle Nahrungsfette aber nur, wenn sie zu den oben genannten Lipolyseprodukten zerlegt wurden. Glycin sowie kurz- und mittelkettige Fettsäuren können auch resorbiert werden, ohne dass sie Mizellen gebildet hätten.
Zur Absorption treten die Mizellen mit den Enterozyten des Dünndarms in Kontakt, zerfallen und geben die gespaltenen Fette frei. Diese werden dann über noch nicht hinreichend geklärte Mechanismen in den Enterozyten aufgenommen. Langkettige Fettsäuren, Cholesterin, Monoglyceride und Lysophospholipide werden wohl überwiegend über Transporter aufgenommen. Glycerin sowie kurz- und mittelkettige Fettsäuren können dagegen frei in die Enterozyten diffundieren. Im Zellplasma der Enterozyten werden die Fettsäuren durch Übertragung auf Coenzym A aktiviert und zusammen mit den anderen Lipolyseprodukten zum glatten ER transportiert, wo die Triacylglyceride erneut gebildet, Cholesterin wieder verestert und Lysophospholipide wieder in Phospholipide umgewandelt werden. Die Triglyceride, Cholesterinester und Phospholipide verbinden sich dann mit fettlöslichen Vitaminen und Apolipoproteinen aus dem rauen ER zu Chylomikronen. Diese werden dann an die Lymphe abgegeben, von wo aus sie in den Milchbrustgang und letztlich im linken Venenwinkel ins Blut gelangen. Glycerin und die kurz- und mittelkettigen Fettsäuren gelangen vom Enterozyten direkt ins Blut und weiter in die Leber.
Aufnahme von Vitaminen
Alle fettlöslichen Vitamine (Vitamin A, Vitamin D3, Vitamin E und Vitamin K) werden zusammen mit den Fetten in den Mizellen absorbiert (siehe Aufnahme der Fette). Für die wasserlöslichen Vitamine gibt es spezifische Transportproteine:
- Vitamin C, Vitamin H (Biotin) und Vitamin B5 (Pantothensäure) gelangen zusammen mit Natrium über einen Natrium-Symporter in die Enterozyten.
- Vitamin B3 (Niacin) wird über einen H+-Symporter aufgenommen.
- Auch für die Vitamine B1 und B2 existieren Transportproteine.
- Vitamin B6 tritt über einen Kanal (erleichterte Diffusion) in die Enterozyten.
- Vitamin B9 (Folsäure) wird von Folat-Polyglutamat als Folat-Monoglutamat hydrolytisch abgespalten und durch einen Folat-Monoglutamat/OH−-Antiporter absorbiert. Die Hydrolyse von Folat-Polyglutamat katalysieren die Bürstensaumpeptidasen.
- Vitamin B12: Im Ileum wird mithilfe des aus dem Magen stammenden intrinsischen Faktors Vitamin B12 (Cobalamine) durch rezeptorvermittelte Endozytose aufgenommen. Dazu wird es an den intrinsischen Faktor gekoppelt.
Aufnahme von Wasser
Täglich werden im Dünndarm zirka sechs Liter Flüssigkeit resorbiert (von ursprünglichen acht Litern, die in den Dünndarm gelangen). Der Dickdarm absorbiert nochmals etwa zwei Liter Wasser, so dass nur ca. 100 ml am Tag ausgeschieden werden. Die maximal mögliche Wasseraufnahme des Dünndarms liegt bei etwa 15 bis 20 Litern Wasser pro Tag. Die Resorption selbst erfolgt parazellulär und transzellulär und ist an den Transport wasserlöslicher Substanzen (wie z. B. Natrium) gebunden. Das Wasser folgt quasi dem osmotischen Druckgradienten. Die Menge an absorbierten Wasser sinkt Richtung Dickdarm kontinuierlich ab. Grund ist die zunehmende Dichtheit der Zellverbindungen untereinander (der Tight junctions).
Aufnahme von Mineralstoffen
-
Natrium: Von den etwa 30 g Natrium am Tag, die in den Darm gelangen (25 g aus Verdauungssäften und 5 g aus der Nahrung) werden ca. 75 % im Dünndarm und 24 % im Dickdarm resorbiert; nur etwa 1 % geht verloren. Die Absorption erfolgt sowohl para- als auch transzellulär:
- Natrium wird zusammen mit Kohlenhydraten oder Aminosäuren aufgenommen (siehe Abschnitte weiter oben)
- Im Zwölffingerdarm und Leerdarm finden sich Na+-H+-Antiporter (wie der NH3-Transporter), die Natrium in die Zelle und ein Proton ins Dünndarmlumen transportieren. Diese Transporter werden von Hydrogencarbonat (Bikarbonat) stimuliert, das besonders in den Sekreten der Galle, Bauchspeicheldrüse und des Dünndarms selbst enthalten ist.
- Zwischen den Verdauungsphasen gewinnt der Transport durch parallel arbeitende Na+/H+- und Cl−/HCO3−-Antiporter im Krummdarm an Bedeutung. Diese Transportsysteme stellen hier den quantitativ größten Anteil.
- Parazellulärer Transport: Im Zwölffingerdarm und Leerdarm sind die Verbindungen zwischen den Zellen relativ schwach ausgeprägt und Wasser kann einfach zwischen den Zellen hindurchfließen. Dabei reißt es kleinere gelöste Moleküle mit (Solvent drag)
- Chlorid wird im Zwölffingerdarm und Leerdarm parazellulär (wie Natrium) transportiert. Im Krummdarm erfolgt der Transport hauptsächlich über den genannten Cl−/HCO3−-Antiporter.
- Kalium wird im Leerdarm und Krummdarm parazellulär absorbiert.
- Calcium: Täglich wird ca. 1 g Calcium aufgenommen, gleichzeitig werden aber mit den Verdauungssekreten etwa 325 mg abgegeben. Bei einer Aufnahme von etwa 500 mg Calcium beträgt der tatsächliche Zuwachs an Calcium also nur 175 mg am Tag und der Großteil wird mit dem Stuhl wieder ausgeschieden. Im Zwölffingerdarm gibt es einen aktiven Ca2+-Transporter, der Calcium in die Zelle pumpt, wo es an Calbindin gebunden und anschließend über einen 3Na+/Ca2+-Antiporter oder eine Ca2+-ATPase ans Blut abgegeben wird. Diese aktive Form der Aufnahme wird durch Vitamin D3 stimuliert. Der Großteil des Calcium wird jedoch passiv, also parazellulär, im Krummdarm und Leerdarm resorbiert.
- Magnesium wird im Zwölffingerdarm passiv und im Leerdarm aktiv resorbiert. Insgesamt werden von den etwa täglich zugeführten 300–400 mg etwa 30–40 % aufgenommen und der Rest ausgeschieden. Ein Teil des Magnesiums stammt, ähnlich wie Calcium, von den Verdauungssäften.
- Phosphat: Organische Phosphate müssen zunächst hydrolysiert werden, bevor sie aufgenommen werden können. Dann werden sie wie die ca. 1 g anorganisches Phosphat über einen Na+-Symporter aufgenommen. Die Absorption wird ebenfalls über Vitamin D3 stimuliert.
- Sulfat wird im Leerdarm und Krummdarm als anorganisches Sulfat (SO42−) über einen Na+-Symporter aufgenommen. Daneben scheint es noch andere relevante Sulfat-Transporter zu geben. Ansonsten wird Schwefel als Bestandteil der Aminosäuren Cystein und Methionin absorbiert.
- Kupfer wird im Magen und im Zwölffingerdarm resorbiert. Die gesamte Resorption beträgt etwa 10 % des in der Nahrung enthaltenen Kupfers. Der genaue Absorptionsmechanismus ist nicht bekannt. Anschließend wird es ans Blut abgegeben und an Albumin und Transcuprein gebunden und erreicht so die Leber.
- Zink wird im Leer- und Krummdarm aufgenommen. Die Resorption beträgt etwa 10–40 %. Auch Zink wird im Blut an Albumin gebunden.
- Auch Mangan wird im Dünndarm aufgenommen und dann an β1-Globulin im Blut transportiert.
- Cobalt: Zwar beträgt die Resorption 70–100 %, aber es wird mit dem Urin schnell wieder ausgeschieden.
- Die Absorption von Fluorid ist ähnlich effektiv wie die von Cobalt. Es wird fast ausschließlich in Knochen und Zähne eingebaut und ansonsten mit dem Urin wieder ausgeschieden.
- Auch Selen, Molybdän und Chrom werden vom Körper benötigt. Selen wird zwischen 50 und 100 % resorbiert.
- Obwohl sie keine Funktion erfüllen und in höherer Dosis giftig sind, nimmt der Körper auch Cadmium, Blei und Quecksilber auf, da diese in der Nahrung vorkommen.
Aufnahme von Eisen
Die Eisenresorption ist mit etwa 10–20 % relativ ineffizient. Täglich werden durchschnittlich 10–15 mg Eisen mit der Nahrung aufgenommen und somit nur 1 bis 1,5 mg absorbiert. Bei erhöhten Bedarf (z. B. bei einer Schwangerschaft) kann die Effizienz jedoch auf bis zu 40 % steigen. Es existieren zwei Mechanismen:
- Freies Eisen kann nur als Fe2+ aufgenommen werden. Daher wird Fe3+ zuerst durch die Ferrireduktase und mittels freier SH-Gruppen und Vitamin C zu Fe2+ reduziert. Einige Substanzen wie Gerbsäure (Tee), Phosphate (z. B. im Eigelb) u. a. hemmen die Resorption. Im Zwölffingerdarm gelangt das Fe2+ dann über den DMT1-Transporter zusammen mit einem Proton in die Zelle.
- Häm-gebundenes Eisen wird vom heme carrier protein 1 vom Protein abgespalten und in die Zelle transportiert. Dort wird es von der Hämoxigenase zu Fe2+ reduziert und anschließend Fe2+ aus dem Häm freigesetzt. Somit liegt nun ebenso wie beim freien Eisen freies Fe2+ in der Zelle vor.
Das Fe2+ wird in beiden Fällen an Mobilferrin gebunden, das es zur basolateralen Seite der Zelle transportiert. Auf dieser Seite hat die Zelle Anschluss zum Blut. Fe2+ wird dort von Hephaestin wieder zu Fe3+ oxidiert, welches durch den Transporter IREG ans Blut übergeben wird. Hephaestin und IREG binden dabei einen gemeinsamen Komplex. Im Blut bindet Fe3+ an Apotransferrin – das Transportprotein für Eisen im Blut.
In der Mukosazelle kann Eisen aber auch als Ferritin gespeichert werden und wird dann beim Abfallen der Zelle ins Darmlumen (Desquamation) ausgeschieden. Die Ausscheidung ist mit 1–2 mg pro Tag jedoch sehr gering und daher kann es bei übermäßiger Resorption auch zu einer Eisenüberdosierung kommen; ein Eisenmangel ist aber wesentlich weiter verbreitet.
Untersuchungsmöglichkeiten des Dünndarmes
- Anamnese (Beschwerden erfragen)
- Auskultation (Darm abhören), Palpation (Bauch abtasten)
- Ultraschall des Dünndarmes, des Mesenteriums und seiner Gefäße
- Biopsie aus dem Zwölffingerdarm
-
Blutuntersuchungen
- Entzündungszeichen
- Resorptionsmangelzeichen
- Stuhluntersuchungen
- auf Keime
- auf Chymotrypsin
- auf Stuhlfettgehalt
- Röntgen – Abdomenübersicht (des Bauches)
- Röntgen mit Kontrastmittelpassage (zur besseren Darstellung des Darms)
- Röntgen – Dünndarmdoppelkontrastuntersuchung nach Sellink
- CT oder MRT des Abdomens (Bauches)
- Spiegelung des Zwölffingerdarmes
- Kapselendoskopie
- Koloskopie des terminalen Ileums (Ileoskopie)
- orale Push-Enteroskopie
- Doppel- oder Einzelballon-Enteroskopie des Dünndarmes (Push-and-Pull)
- Dünndarmmanometrie
Krankheiten des Dünndarms
- Morbus Crohn
- Meckel-Divertikel
- Ulcus duodeni (Zwölffingerdarmgeschwür)
- Dünndarmatresie
- bakterielle Dünndarmfehlbesiedlung, in 40 Prozent der Fälle vergesellschaftet mit einer exokrinen Pankreasinsuffizienz (EPI)
- Morbus Whipple
- Volvulus
- Invagination
- Duodenaldivertikel
- Nahrungsmittelunverträglichkeiten (z. B. Zöliakie, Laktoseintoleranz, Weizenallergie)
- Pilze (z. B. Candida albicans)
- Parasiten
Verletzungen des Dünndarms wurden von Hippokrates von Kos (um 350 v. Chr.) und bei Aulus Cornelius Celsus (um 20 n. Chr.) noch für unheilbar gehalten. Gabriele Falloppio wandte sich im 16. Jahrhundert gegen die Lehre von der Unheilbarkeit der Dünndarmwunden.
Krankheitsanzeichen
Es gibt verschiedene Symptome, die bei Erkrankungen des Dünndarms auftreten können. Verschiedene Erkrankungen können über unterschiedliche Mechanismen Durchfall verursachen. Auch Verstopfung kann verschiedene Ursachen haben. Hinter einem Ileus, im Deutschen als Darmverschluss bezeichnet, kann sich entweder ein tatsächliches mechanisches Hindernis (mechanischer Ileus) oder eine Lähmung des Darmes (funktioneller Ileus) verbergen. Blutungen des Dünndarms entstehen meistens auf dem Boden entzündlicher Prozesse und aus Geschwüren. Weitere unspezifische Krankheitsanzeichen sind Bauchschmerzen, die in der Regel nicht auf einen bestimmten Darmabschnitt zu lokalisieren sind, sowie ungewollter Gewichtsverlust und Mangelernährung.
Tumorerkrankungen
Tumoren des Dünndarms sind beim Menschen selten, sie machen nur etwa 3 % aller Tumoren der Verdauungsorgane aus. Es gibt keine charakteristischen Symptome: die Tumoren können krampfartige Bauchschmerzen hervorrufen, Blut absondern oder durch ihr Wachstum das Lumen des Dünndarms verengen. Die meisten dieser Tumoren sind gutartig. Sie gehen meistens vom Drüsenepithel aus, sie sind damit Adenome, die auch Polypen bilden können. Daneben finden sich Leiomyome (Tumoren der glatten Muskelzellen), Lipome (Tumoren der Fettzellen) und Angiome. Bösartiger Krebs ist sehr selten, findet sich aber gehäuft bei Patienten mit Morbus Crohn, Zöliakie oder AIDS. Auch die bösartigen Tumoren gehen meistens von den Drüsenzellen aus (Adenokarzinome), diese Gruppe macht etwa die Hälfte der bösartigen Dünndarmtumoren aus. Etwa jeder fünfte bösartige Tumor ist die Manifestation eines Lymphoms. Daneben gibt es Karzinoidtumoren, die am häufigsten im distalen Ileum zu finden sind, sowie Gastrointestinale Stromatumoren und Leiomyosarkome. Papillentumoren betreffen zwar den Dünndarm, sind aber keine eigentlichen Dünndarmtumoren, da sie vom Gallen- bzw. Pancreasgang ausgehen.
Behandlungsmethoden des Dünndarmes
- Ausreichender Flüssigkeits-, Salz- und Zuckerersatz bei Durchfall
- Nahrungskarenz
- parenterale Ernährung
- Spülung mit physiologischer Kochsalzlösung
- Antibiotikumtherapie
- Operation (z. B. Dünndarmresektion), auch bei Dünndarmwunden
- Cortison
- Diät ohne bestimmte Nahrungsbestandteile (Gluten, Laktose etc.)
- Viszerale Osteopathie
Literatur
- Franz-Viktor Salomon: Darm, Intestinum (Enteron). In: Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. Enke-Verlag Stuttgart, 2. erw. Aufl. 2008, S. 293–311, ISBN 978-3-8304-1075-1.
- Gerhard Aumüller et al.: Duale Reihe Anatomie, 2. Auflage. Georg Thieme Verlag, Stuttgart 2010, ISBN 978-3-13-152862-9.
- Giulia Enders; Jill Enders (Illustrationen): Darm mit Charme. Alles über ein unterschätztes Organ. Ullstein, Berlin 2014, ISBN 978-3-550-08041-8 (Taschenbuch) / ISBN 978-3-550-08108-8 (gebunden).
- Michael Schünke et al.: Prometheus – Lernatlas der Anatomie. Innere Organe, 4. Auflage, Georg Thieme Verlag, Stuttgart · New York, 2015, ISBN 978-3-13-139534-4.
- Renate Lüllmann-Rauch: Taschenlehrbuch Histologie. 5. Auflage. Georg Thieme Verlag, Stuttgart 2015, ISBN 978-3-13-129245-2.