
Das Endhirn oder Telencephalon (altgriechisch τέλος telos, deutsch ‚Ende‘ und ἐγκέφαλος enkephalos, deutsch ‚Gehirn‘) ist ein Teil des Gehirns und damit des Zentralnervensystems. Er geht aus dem vorderen Bereich des Vorderhirns hervor und entwickelt sich bei Säugetieren (wie dem Menschen) zum größten der fünf Hirnabschnitte.

Andere gebräuchliche Namen für diesen Hirnabschnitt sind auch Großhirn und Cerebrum (von lateinisch cerebrum ‚Gehirn‘, ‚Hirn‘). Cerebrum bezeichnet fachsprachlich jedoch auch das Gehirn insgesamt.

Anatomie
Anordnung Grauer und Weißer Substanz in Hemisphären
Aus dem unpaaren Hirnbläschen des Endhirns entwickelt sich ein Paar ungefähr halbkugelförmiger Gebilde, die Hemisphärenbläschen, die je zur Großhirnhemisphäre (Hemisphaerium cerebri) einer Seite werden. Neben der unpaaren medianen Gewebebrücke (Lamina terminalis) entstehen beide Hemisphären miteinander verbindende Querbahnen, die Kommissursysteme des Endhirns wie der breite Balken (Corpus callosum). Linke und rechte Hemisphäre werden durch eine tiefe Längsfurche (Fissura longitudinalis cerebri) geschieden und umschließen je einen Seitenventrikel als inneren Liquorraum. Jede der Hemisphären enthält innen gelegene Kerngebiete als Basalganglien (Nuclei basales), die aus dem ventralen (bauchseitigen) Telencephalon entstehen. Diese werden umhüllt vom außen liegenden Hirnmantel (Pallium), der aus dem dorsalen (rückenseitigen) Telencephalon hervorgeht.
Die Oberfläche des Palliums besteht als Rinde des Endhirns oder Großhirnrinde (Cortex cerebri) – ähnlich wie die Kleinhirnrinde (Cortex cerebelli) – aus Grauer Substanz mit Nervenzellkörpern in mehreren Schichten. Darunter liegt als sogenanntes Marklager aus Weißer Substanz ein dichtes Geflecht von Nervenfasern, deren Nervenzellfortsätze vielfältige Verbindungen schaffen. Als Assoziationsfasern verbinden sie unterschiedliche Areale des Cortex cerebri der Hemisphäre gleicher Seite untereinander, als Kommissurfasern mit solchen der Gegenseite und als Fasern von Projektionsbahnen stellen sie auf- oder absteigend Verbindungen von und zu verschiedenen anderen Regionen des Gehirns bzw. des zentralen Nervensystems dar.


Verbindungen beider Großhirnhälften (Commissurae)
Die beiden Hemisphären sind durch Fasersysteme gegenseitig miteinander verbunden, die als Kommissuren (Commissurae) die Medianebene queren; von diesen drei ist der Balken am faserreichsten:
- Corpus callosum (auch Balken genannt)
- Commissura anterior bzw. rostralis
- Commissura fornicis
Basale Kerne (Nuclei basales)
Die aus dem ventralen Telencephalon hervorgehenden subkortikalen Kerne werden auch als basale Kerne oder Nuclei basales bezeichnet bzw. als Basalganglien. Sie liegen an der Basis des Endhirns seitlich und bauchseitig eines Seitenventrikels und schließen an Bereiche des Thalamus des Zwischenhirns. Infolge der hindurchziehenden Fasern von und zur Großhirnrinde hat ein Teil der Kerne ein gestreiftes Aussehen, weshalb diese auch als Corpus striatum (Streifenkörper) bezeichnet werden.
Hirnlappen (Lobi)
Die Großhirnrinde einer Hemisphäre lässt sich jederseits in fünf oder sechs Hirnlappen (Lobi cerebri) einteilen. Vier davon liegen an der Oberfläche (siehe Abbildung rechts), der Insellappen wird vom Operculum verdeckt und auch der Limbische Cortex, von einigen Fachleuten als sechster Hirnlappen (Lobus limbicus) aufgefasst, liegt in der Tiefe.

Stirn-/Frontallappen (Lobus frontalis, gelb)
Scheitel-/Parietallappen (Lobus parietalis, rot)
Hinterhaupts-/Okzipitallappen (Lobus occipitalis, blau)
Schläfen-/Temporallappen (Lobus temporalis, grün)
Insellappen (Insula oder Lobus insularis) – verdeckt
Bei genauerer Betrachtung zeigen die einzelnen Furchen (Sulci; Singular: Sulcus) und Windungen (Gyri; Singular: Gyrus) der verschiedenen Hirnlappen keine exakte Symmetrie im Vergleich von linker und rechter Seite der Großhirnhemisphären eines Individuums. Einige dieser neuroanatomischen Ungleichheiten der im Prinzip bilateral symmetrisch aufgebauten Hälften hängen auch damit zusammen, dass einige der cerebralen Funktionen bevorzugt in einer der beiden Großhirnhälften ausgeführt werden. Diese prozedural unterschiedliche Aufteilung wird auch als Lateralisation bezeichnet.
Der oberflächliche Teil der Hemisphären beziehungsweise ihr Pallium (Hirnmantel) ist – bis auf wenige epithelial bleibende Wandbezirke – ausgebildet als nur wenige Millimeter dicke Hirnrinde, der Cortex cerebri. Das Gewebe dieses Cortex des Großhirns – oft wird der Ausdruck Pallium synonym verwendet – zeigt allerdings histologisch nicht überall den gleichen Aufbau. Weite Bereiche der Rinde sind recht ähnlich aus sechs unterschiedlich starken Schichten (Laminae) aufgebaut. Von diesem sogenannten Isocortex verschieden ist der aus zwei oder drei Schichten anders aufgebaute Allocortex. Auch unter entwicklungsgeschichtlichen Gesichtspunkten ist der Cortex cerebri nicht einheitlich und kann so in die Anteile von Paläo-, Archi- und Neocortex (bzw. -pallium) unterschieden werden.
Entwicklungsgeschichtliche Einteilung
Wie beim Gehirn als ganzem, so bietet sich auch beim Telencephalon eine Unterscheidung seiner Anteile unter phylogenetischer oder embryogenetischer Perspektive an.
Paläocortex
Der Paläocortex (bzw. das Palaeopallium oder Paläopallium) besitzt mit einem primitiven zweischichtigen Aufbau den urtümlichsten Typ einer Hirnrinde, einem „Althirn“ entsprechend. Es handelt sich hierbei um einen entwicklungsgeschichtlichen Begriff. Der Paläocortex liegt am vorderen unteren Teil der Hemisphären. Als Grenze zum Neocortex gilt der Sulcus rhinalis lateralis.
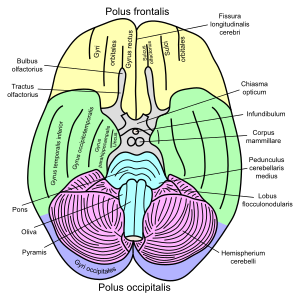
Hirnlappen der Hemisphären farbig dargestellt, ähnlich wie in Abbildung oben.
Beim Menschen wird der Riechkolben (Bulbus olfactorius, grau), im Sulcus olfactorius liegend, vom Stirnlappen überragt. Tractus, Trigonum, Striae olf. und Substantia perforata anterior liegen im grau eingefärbten Bereich noch vor dem Chiasma opticum.
Der Riechkolben (Bulbus olfactorius) erhält über die Nn. olfactorii unmittelbar Afferenzen des Geruchssinnes von den Riechzellen und stellt somit ein primäres Riechgebiet dar. Mitsamt der anschließenden zentralen Bahn und den sekundären olfaktorischen Projektionsgebieten des Paläocortex wird deshalb auch zusammenfassend vom Riechhirn oder Rhinencephalon gesprochen. Hierzu zählen
als primäre Gebiete
- Bulbus olfactorius (Riechkolben)
- mit Pedunculus olfactorius (Riechstiel)
- und folgendem Tractus olfactorius (Riechbahn), der sich
- zum Trigonum olfactorium (Riechdreieck) verbreitert und dann in
- die Striae olfactoriae lateralis et medialis aufzweigt, zwischen denen
als sekundäre Gebiete
- die Substantia perforata anterior jederseits (mit dem Tuberculum olfactorium) liegt;
- weitere Projektionen erreichen auch Gyrus ambiens und Gyrus semilunaris (im Lobus piriformis) auf der medialen Seite des Schläfenlappens als Anteile des Paläocortex
Der Paläocortex wird wegen seiner histologischen Eigentümlichkeiten dem Allocortex zugerechnet (mit anderem Aufbau als der Isocortex).
Archicortex
Der Archicortex (oft synonym: Archipallium) kann entwicklungsgeschichtlich als ein Stadium zwischen Paläocortex und Neocortex angesehen werden. Ein über olfaktorische Projektionsgebiete hinausgehender Teil der Hirnrinde tritt andeutungsweise erstmals bei Reptilien auf und ist histologisch deutlich vom jüngeren Neocortex zu unterscheiden. Mit der späteren Entwicklung des Neocortex wird der Archikortex in seiner Ausdehnung reduziert und auf die Innenseite des Temporallappens verdrängt.
Der Archicortex besteht aus Hippocampus, Gyrus dentatus und Fimbria fornicis und gehört auch zum limbischen System, dem eine Schlüsselrolle in der Verarbeitung emotionaler Inhalte zugeschrieben wird. Diese Formationen des Archicortex gehören zum sogenannten Randbogen. Durch die Kommissurenbahnen des Corpus callosum als Formation des Neocortex wird dieser Randbogen in einen inneren und äußeren Bogen unterteilt. Zum äußeren Randbogen zählt auch der Gyrus cinguli und das Indusium griseum. Letzteres stellt topographisch eine Fortsetzung des Gyrus dentatus dar. Histologisch gehören auch die meisten Areale des Archicortex zum Allocortex; der Gyrus cinguli des äußeren Bogens allerdings zeigt schon den vielschichtigen Aufbau des Isocortex.
Neocortex
Der Neocortex (oft synonym: Neopallium) ist der entwicklungsgeschichtlich jüngste und am meisten differenzierte Teil des Gehirns. Unter Neocortex kann man rund neun Zehntel der Großhirnrinde (Cortex cerebri) verstehen. Beide Bezeichnungen sind aber nicht identisch (siehe dazu auch die methodische Differenzierung der verschiedenen Begriffe und die histologische Zugehörigkeit des Neocortex zum Isocortex).