
| Marburg-Virus | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

Marburg-Virus | ||||||||||||||||||||
| Systematik | ||||||||||||||||||||
| ||||||||||||||||||||
| Taxonomische Merkmale | ||||||||||||||||||||
| ||||||||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||||||||
| Marburg marburgvirus | ||||||||||||||||||||
| Kurzbezeichnung | ||||||||||||||||||||
| MARV | ||||||||||||||||||||
| Links | ||||||||||||||||||||
|
Das Marburg-Virus ist ein behülltes Einzel(−)-Strang-RNA-Virus (ss(−)RNA) der Familie Filoviridae und Gattung Marburgvirus und der Erreger des Marburgfiebers. Erkrankungen von Menschen wurden erstmals 1967 im hessischen Marburg festgestellt, und ihr Erreger wurde im selben Jahr in Hamburg als ein neues Virus identifiziert. Seither trat das Marburg-Fieber mehrfach in aller Welt auf. Seine Letalität liegt bei mindestens 23 Prozent.
Größe und Gestalt
Dieses Virus besitzt eine fadenförmige (lateinisch filum, „Faden“) Gestalt, diese Grundstruktur kann langgestreckt vorliegen oder gekrümmt bzw. gebogen sein. Dabei ergeben sich Strukturen, die U-förmig, kreisförmig, wie die Ziffer 6 oder wie ein Angelhaken gebogen sind. Das Virus hat eine Länge von etwa 800 nm (Bereich von 130 bis 2600 Nanometer), während der Durchmesser relativ konstant bei 80–100 nm liegt. Im elektronenmikroskopischen Bild lassen sich kreuzförmige Riefen erkennen. Untersuchungen von 2011 mit Hilfe der Kryoelektronentomographie ergeben eine durchschnittliche Länge von 892 nm, der Durchmesser liegt bei durchschnittlich 91 nm. Zusammen mit den Ebolaviren aus derselben Familie gehört das Marburg-Virus zu den größten bekannten RNA-Viren.
Aufbau des Virions
Das Virion besteht aus einer Virushülle, die das helikale Kapsid umgibt, in dem sich das Virusgenom befindet. Am Aufbau des Virions sind insgesamt sieben Strukturproteine beteiligt. In infizierten Wirtszellen erfolgt die Replikation innerhalb von intrazellulären Einschlüssen, in denen Vorstufen des Nukleokapsids gebildet werden. Diese röhrenförmigen Strukturen mit einem äußeren Durchmesser von 45–50 nm werden durch die Nukleoproteine (NP) gebildet. Zusammen mit den viralen Proteinen VP30, VP35 und L entsteht hieraus das Nukleokapsid. Das virale Genom ist am NP gebunden, weshalb man von einem Nukleokapsid spricht. Das NP ist notwendig für die Transkription und Replikation und ist ebenfalls beim sogenannten budding, dem Abschnüren von Viruspartikeln aus der Wirtszelle, von Bedeutung. Der Komplex aus den Proteinen VP35 und L dient als RNA-abhängige RNA-Polymerase, wobei VP35 den Cofaktor der Polymerase darstellt. Neben NP, VP35 und L ist auch VP30 an der Transkription und Replikation beteiligt, allerdings ist bei diesem Protein die genaue Wirkungsweise im Ablauf noch nicht geklärt.
Die beiden Matrixproteine VP40 (Hauptkomponente) und VP24 stellen die Verbindung der Virushülle mit dem Nukleokapsid dar, wobei im Querschnitt des Virions VP24 eher im Inneren und VP40 im äußeren Teil vorhanden ist. VP40 kann leicht vom Nukleokapsid abgespalten werden. Nach der Proteinsynthese in der Wirtszelle assoziiert VP40 mit der Zellmembran und ist somit beim budding beteiligt, indem Teile der Zellmembran die Virushülle bilden. Obwohl VP24 eher im Inneren des Virions lokalisiert ist, lässt sich dieses virale Protein durch Salzlösungen mit ansteigender Konzentration vom Nukleokapsid ablösen. Bei der Freisetzung von Viruspartikeln ist VP24 im Anschluss an die Replikation und vor dem budding beteiligt. Die Virushülle enthält neben den Lipiden der Zellmembran, die von der Wirtszelle stammen, noch virale Glykoproteine (GP). Beim GP handelt es sich um ein Transmembranprotein, das in der Virushülle verankert ist und zur Außenseite stachelförmige Ausbuchtungen (englisch spikes) bildet. In der Wirtszelle wird es während des Transports vom endoplasmatischen Retikulum zur Zellmembran glykosyliert, d. h., es werden Kohlenhydratketten an das Protein angelagert. Die Funktionen von GP liegen in der Anhaftung an die Wirtszelle an spezielle Rezeptoren und in der Fusion der Virushülle.
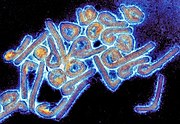
Abb. 2: kolorierte EM-Aufnahne von Virionen des Marburg-Virus.

Abb. 3: Modell des Marburg-Virus. Maßstab 5 nm.

Abb. 4: (a) RTEM- und (b) TEM-Aufnahmen von Marburg-Viruspartikeln, aus infizierten Vero-E6-Zellen knospend.




Abbildung 3 zeigt einen Querschnitt des Virions in der Kryoelektronentomographie (Kryo-ET). Im Inneren ist die RNA (rot) zu erkennen, verbunden mit den Nukleoproteinen (gelb) und weiteren viralen Proteinen, die gemeinsam das Nukleokapsid bilden. Dieses ist umgeben von der Virushülle (grün bis blau), deren Glykoproteine als etwa 10 nm lange Vorsprünge (dunkelblau) zu erkennen sind.
Genom
Das virale Genom liegt in Form eines einzelsträngigen RNA-Moleküls (Ribonukleinsäure) vor, mit (–)-Polarität (antisense). Die RNA umfasst 19 kb (Kilo-Basenpaare), wobei bei einem Einzelstrang keine Basenpaare, sondern ungepaarte Basen in den Nukleotiden vorkommen. Genetische Analysen zeigen, dass die RNA aus 19.111 bis 19.114 Nukleotiden aufgebaut ist. Das Genom umfasst sieben Gene, die als Cistrone gelten, und linear hintereinander liegen. Jedes Gen besteht aus einem hoch konservierten Start- und Stoppsignal, einem ungewöhnlich langen Genabschnitt am 3'- und 5'-Ende, der nicht für die Translation verwendet wird, und dem dazwischenliegenden offenen Leserahmen (engl. open reading frame, ORF). Die Gene können durch kurze Abschnitte nichtcodierender Ribonukleinsäure (engl. intergenic regions, IR) voneinander getrennt sein, wie dies beispielsweise bei den Genen für GP und VP30 der Fall ist. Die IR-Abschnitte umfassen 4 bis 97 Nukleotide. Alternativ können sich das Stoppsignal und das Startsignal des nächsten Gens überlappen; dies trifft beispielsweise bei den Genen für VP30 und VP24 zu. Diese Überlappung ist nur bei Vertretern der Familie der Filoviridae zu finden. Am 3'- und am 5'-Ende befinden sich Abschnitte, die als Promotor für die Transkription und die Replikation dienen.
Abbildung 6 zeigt den prinzipiellen Aufbau des Genoms des Marburg-Virus. Vom 3'- zum 5'-Ende liegen linear angeordnet die Gene, die für die sieben viralen Proteine NP, VP35, VP40, GP, VP30, VP24 und L codieren. Deren ORF sind unterschiedlich eingefärbt. Abschnitte, die zu einem Gen gehören, aber nicht für die Translation verwendet werden, sind als hellgraue Kästchen dargestellt, die IR sind als dunkelgraue Verbindungsstücke angegeben und die Anfangs- und Endsequenz am 3'- bzw. 5'-Ende erscheinen schwarz. Grüne Dreiecke symbolisieren die Startsignale für die Transkription, während rote Streifen die Stoppsignale darstellen.
Reservoir und Verbreitung
Das Reservoir, aus dem das Virus stammt, bzw. sein Reservoirwirt ist bis heute nicht genau bekannt. Vermutlich ist der Überträger der Nilflughund, eine Flughundart, die in Europa und Afrika vorkommt. Das Virus und virusspezifische Antikörper konnten in ihrem Blut nachgewiesen werden, auch in Regionen, in denen bislang keine Erkrankungsfälle registriert wurden. Da der Flughund sein Quartier in Höhlen hat, kann man Ausbrüche unter Bergleuten mit ihm in Verbindung bringen.
Das Marburg-Virus stammt primär aus Afrika und kommt in den Ländern Uganda, Kenia (West-Kenia) und vermutlich Simbabwe vor. Es kam offenbar über infizierte Affen von Uganda nach Hessen, wo die Primaten bei Tierversuchen eines Pharmakonzerns eingesetzt werden sollten. Eine weitere Ausdehnung wird von Wissenschaftlern für wahrscheinlich gehalten. In Europa wurden 1967 die ersten Erkrankungsfälle dokumentiert. 2018 und 2020 wurde das Virus auch in Westafrika bei Flughunden in Sierra Leone nachgewiesen. Im August 2021 wurde das Virus erstmals bei einem einzelnen Patienten in Guinea nachgewiesen, sodass es sich hierbei um die erste belegte Übertragung auf den Menschen in Westafrika handelt.
Virulenz
Bei diesem Virus handelt es sich um einen hochpathogenen Erreger, der beim Menschen das Marburgfieber, ein hämorrhagisches Fieber, auslöst. Die Sterblichkeit bei dieser Erkrankung liegt laut den Centers for Disease Control and Prevention (CDC) bei mindestens 23 bis 25 Prozent. Bei Ausbrüchen im Kongo und in Angola lag sie jedoch wesentlich höher (siehe Krankheitsfälle). Diese hohe Sterblichkeit deutet – wie bei den Ebolaviren – darauf hin, dass weder das Virus an den Menschen noch der Mensch an das Marburg-Virus angepasst ist, da es vor allem andere Wirte infiziert. Die rasche Schädigung seines Wirtes bis hin zu seinem Tod ist zudem für ein Virus kein vorteilhafter Effekt, da es zur eigenen Vermehrung und Verbreitung auf einen lebenden Wirt angewiesen ist.
Übertragung
Das Marburg-Virus wird durch den Austausch von Körperflüssigkeiten und durch Schmierinfektion bzw. Kontaktinfektion übertragen. Viruspartikel bleiben innerhalb von geronnenem Blut für einen Zeitraum von 4–5 Tagen infektiös. Bei Rekonvaleszenten kann das Virus auch nach Ablauf der akuten Infektion für einige Monate an einzelnen Stellen des Körpers – insbesondere dem Sperma – in pathogener Form zurückbleiben.
Systematik

Die Gattungen Ebolavirus und Marburgvirus in der Familie der Filoviridae sind eng miteinander verwandt, was sich unter anderem in der Struktur der Virionen zeigt. Die Viruspartikel der Marburg-Viren sind allerdings deutlich kleiner als die der Ebolaviren, wobei das Genom des Marburg-Virus (MARV) etwas größer ist als das des Ebola-Virus (EBOV).
In der Gattung Marburgvirus ist lediglich eine Spezies enthalten, die früher als Lake Victoria Marburg-Virus bezeichnet wurde. Die ICTV Filoviridae Study Group hat 2010 eine aktualisierte Systematik und Nomenklatur der Vertreter der Filoviridae vorgeschlagen, die in dem 9th ICTV Report von 2011 umgesetzt wurde. Demnach soll die folgende Nomenklatur verwendet werden, bei der die Gattung als Marburgvirus und die Spezies als Marburg-Marburgvirus (englisch Marburg marburgvirus) bezeichnet wird.
- Gattung Marburgvirus
- Spezies Marburg-Marburgvirus (en. Marburg marburgvirus, veraltet Lake Victoria Marburg-Virus)
- Virus 1: Marburg-Virus (en. Marburg virus, MARV)
- Virus 2: Ravn-Virus (en. Ravn virus, RAVV)
- Spezies Marburg-Marburgvirus (en. Marburg marburgvirus, veraltet Lake Victoria Marburg-Virus)
Geschichte
Die von dem Virus verursachte Erkrankung trat erstmalig im Jahr 1967 bei Laborangestellten in Marburg (Hessen), später in Frankfurt am Main und Belgrad in Erscheinung. Als am 25. August 1967 mehrere Personen in Marburg starben, wurde die Stadt in eine Art Ausnahmezustand versetzt. Alle Infizierten, auch die später Verstorbenen, hatten zuvor sehr hohes Fieber, gefolgt von Übelkeit, Erbrechen und Durchfall. Diese Symptome deuteten auf eine Infektion mit Salmonellen oder Shigellen hin, diese Bakterien ließen sich aber nicht nachweisen. In der zweiten Woche nach Beginn der Erkrankung kamen hämorrhagische Symptome hinzu, die Patienten bluteten aus Körperöffnungen und nach Nadeleinstichen, von den Blutungen waren auch die inneren Organe, beispielsweise die Leber betroffen. Dies führte bei etwa 25 % der Patienten zum Tode. Als möglicher Krankheitserreger wurde zunächst das Gelbfieber-Virus in Betracht gezogen, konnte aber später ausgeschlossen werden. Auch Bakterien der Gattung Leptospira oder Rickettsia wurden im Verlauf der Untersuchungen ausgeschlossen.
Um die Ätiologie der bis dahin unbekannten Krankheit zu klären, reisten Spezialisten für Tropenkrankheiten im Auftrag der Weltgesundheitsorganisation (WHO) an, darunter Werner Mohr aus Hamburg und der Virologe George B. Dick aus London. Insgesamt acht Laboratorien weltweit untersuchten Blut- und Gewebeproben. In Marburg gelang es nicht, den Erreger auf Zellkulturen zur Vermehrung zu bringen. Da bei den Erkrankten ein sehr hoher Titer von Antikörpern gegen Leptospiren festgestellt worden war, der allerdings, wie sich erst später herausstellte, von einer für Menschen harmlosen Variante stammte, wurde die Züchtung, wie üblich, auf Meerschweinchen versucht. Durch Infektion mit Patientenblut entwickelten die Tiere hohes Fieber, und nach mehrfachen Passagen des Blutes von Tier zu Tier war der Erreger innerhalb von etwa drei Monaten so weit adaptiert, dass keiner der Probanden mehr überlebte. Während dieser Zeit wurden Gewebeteile der inneren Organe neben der lichtmikroskopischen Kontrolle für die elektronenmikroskopische Bearbeitung nach Hamburg verschickt. Hinweise auf die Natur des Erregers ergaben sich jedoch nicht. Daraufhin schlug der Leiter der Abteilung für Virusforschung in Hamburg Dietrich Peters vor, die Suche nach dem Erreger auf das Blut der Meerschweinchen zu richten. Für die Versendung wurden die Proben zur Inaktivierung und Fixierung in mit Formalin gefüllten Gefäßen nach Hamburg geliefert. Nach der Ankunft wurde das Plasma des Blutes, mit einer für die Elektronenmikroskopie von Viren selbst entwickelten Technik, zur Negativkontrastierung (engl. negative stain) vorbereitet. Am 17. November 1967 ließ sich so unmittelbar das gesuchte Pathogen im Elektronenmikroskop als ein bis dahin unbekanntes Virus identifizieren. Als dann wenige Tage später auch im Blut von Patienten dieselben Strukturen im Elektronenmikroskop sichtbar wurden, gab es keinen Zweifel mehr, dass es sich dabei um den Erreger des Marburg-Fiebers handelte. Da er keinen Platz in der aktuellen Systematik fand, wurde mit ihm als Marburg-Virus die neue Familie der Filoviridae begründet, zu der sich einige Jahre später das Ebola-Virus hinzugesellte. An der Untersuchung und der folgenden Veröffentlichung waren Werner Slenczka, Rudolf Siegert und ihr chinesischer Kollege Hsin Lu Shu am Institut für Virologie Marburg sowie Dietrich Peters und Günther Müller am Bernhard-Nocht-Institut für Tropenmedizin beteiligt.
Ein Konsilium der Mediziner Rudolf Siegert, Walter Hennessen und Gustav Adolf Martini gab täglich einen Bericht zur Erforschung des Virus ab. Bis Ende August 1967 starben zwei Tierpfleger und zwei Laborangestellte. 24 Erkrankte lagen im Universitätsklinikum Frankfurt am Main und in der Universitätsklinik der Philipps-Universität Marburg auf der Isolierstation. Insgesamt starben später fünf Menschen in Marburg und zwei in Frankfurt an dem neuen Virus.
Neben der Suche nach dem neuen Virus begann zeitgleich eine epidemiologische Untersuchung, um seine Herkunft zu klären. Das Virus ist höchstwahrscheinlich von infizierten Versuchsaffen – es handelt sich um die Art Äthiopische Grünmeerkatze (Chlorocebus aethiops) – aus Uganda in die Labore des Pharmakonzerns Behringwerke im hessischen Marburg eingeschleppt worden. Deshalb erhielt es auch den Namen Marburg-Virus. Der Pharmakonzern nutzte die Tiere zur Gewinnung von Masern- und Poliomyelitis-Impfstoff. Am Paul-Ehrlich-Institut in der Nähe von Frankfurt am Main wurden diese Impfstoffe geprüft, am Torlak-Institut in Belgrad wurden ebenfalls Impfstoffe hergestellt. Im Nachhinein konnte geklärt werden, dass alle primär Infizierten Kontakt mit Blut, Organen oder Zellkulturen der Äthiopischen Grünmeerkatzen hatten. Informationen über den Gesundheitszustand der Affen ließen sich jedoch nicht ausreichend ermitteln. Wegen des Sechstagekriegs im Juni 1967 konnten die Tiere nicht direkt von Uganda nach Frankfurt transportiert werden, sondern mussten einige Zeit in einem Gehege an einem Londoner Flughafen untergebracht werden. Dabei hatten sie Kontakt zu Finken aus Südafrika und Languren aus Ceylon (heute Sri Lanka). Eine Übertragung des Virus von einer Tierart zur anderen wäre somit theoretisch möglich gewesen, konnte aber weder bewiesen noch ausgeschlossen werden. Die Affen der Art Chlorocebus aethiops wurden in mehreren Lieferungen an die Institute verteilt. In Marburg und Frankfurt wurden keine Auffälligkeiten bezüglich ihres Gesundheitszustandes notiert, allerdings wurden die Versuchstiere nach kurzer Zeit planmäßig getötet. In Belgrad wurden die Tiere noch für sechs Wochen gehalten, dabei wurde eine überdurchschnittlich hohe Mortalitätsrate von 33 % festgestellt. Im Nachhinein ließ sich jedoch nicht mehr klären, ob die Virusinfektion dafür verantwortlich war. In Deutschland wurden die Affen der Art Chlorocebus aethiops, die als Überträger des Virus verdächtig waren, getötet. Dies betraf auch Tiere aus vorhergehenden Lieferungen, die Zahl der insgesamt mit Blausäure getöteten Versuchstiere lag bei über 600.
Der sozialistischen Propaganda diente die zunächst unerklärliche Infektion als Anlass für antiwestliche Propaganda: So behauptete die damalige DDR-Staatszeitung Neues Deutschland, afrikanische Affen seien nur ein Sündenbock, um Geheimversuche bei der Entwicklung von Chemiewaffen zu vertuschen.
Risikogruppe, behördliche Einstufung
Nach der Biostoffverordnung in Verbindung mit der TRBA (Technische Regeln für Biologische Arbeitsstoffe) 462 wird die Virusspezies Marburg Marburg-Virus mit dem Vertreter Marburg-Virus (MARV) in die höchste Risikogruppe 4 eingeordnet.
Durch die Biostoffverordnung werden für biologische Arbeitsstoffe vier Risikogruppen definiert. Das Arbeiten hat unter Berücksichtigung entsprechender Schutzmaßnahmen zu erfolgen, die durch die Biostoffverordnung in vier biologische Schutzstufen (englisch: biosafety level, BSL) eingeteilt werden. Demzufolge müssen Arbeiten mit dem Marburg-Virus unter den strengen Vorgaben der Schutzstufe 4 erfolgen. Weltweit werden Filoviren an 20 Laboratorien erforscht (Stand 2013). Diese müssen daher die Schutzstufe 4 beachten und werden auch als BSL-4-Laboratorien bezeichnet. Für Deutschland trifft dies auf das Bernhard-Nocht-Institut für Tropenmedizin in Hamburg, das Robert Koch-Institut in Berlin, das Friedrich-Loeffler-Institut auf der zum Stadtgebiet von Greifswald gehörenden Insel Riems sowie das Institut für Virologie der Universität Marburg zu.
Verwendung als biologische Waffe
Das Marburg-Virus wurde vom US-amerikanischen CDC als potentieller biologischer Kampfstoff der höchsten Gefahrenklasse eingestuft. Militärisch wurden die Möglichkeiten als biologischer Kampfstoff vom sowjetischen Kampfstoffprogramm Biopreparat erforscht, das 1967 Proben des Virus während des initialen Ausbruchs erlangte. Soweit bekannt, wurden in Bezug auf das Marburg-Virus insbesondere die Verteilung (Distribution) als Aerosol und die Stabilität gefriergetrockneter Viruspartikel erforscht.
Mit einem simulierten Bioterrorismusangriff wurde 1998 ermittelt, dass eine Variola-Marburg-Chimäre einen ökonomischen Schaden von etwa 26 Milliarden US-Dollar pro 100.000 Infizierten bewirken würde.
Krankheitsfälle
- 1967: Marburg, Frankfurt am Main, Belgrad: 31 Infizierte, 7 Tote
- 1975: Johannesburg, Südafrika: 3 Infizierte, 1 Toter
- 1980 Nairobi: 2 Infizierte, 1 Toter (Ursprung: vermutlich Kitum Cave in West-Kenia)
- 1987 Nairobi versterben 2 Rucksacktouristen nach dem Besuch des Mount Elgon National Park.
- In der Stadt Kolzowo bei Nowosibirsk in der Sowjetunion kam es 1988 und 1990 zu insgesamt drei Infektionen. Ein Mensch verstarb.
- 1998 bis 2000: Demokratische Republik Kongo: 149 Infizierte, 123 Tote
- Oktober 2004 bis Mai 2005: Angola, Beginn in der Provinz Uíge: 388 Infizierte, 324 Tote
- Am 21. März 2005 wurde das Marburg-Virus in mehreren Blutproben von Todesopfern in Angola entdeckt. Im April war die Krankheit in sieben Provinzen ausgebrochen. Über 215 Angolaner starben bis dahin bereits am Marburg-Virus. Die meisten Opfer waren jünger als fünf Jahre. Besonders problematisch war die Weigerung der Bevölkerung, die Infizierten zu isolieren. Außerdem gehört traditionell bei den Familien zur Bestattung Verstorbener der persönliche Abschied in Form von Umarmung des Toten und anschließend weitere direkte, persönliche Kontakte der Trauergäste untereinander. Darum war es extrem schwierig, die eigentlich sofort notwendige, unverzügliche Beerdigung ohne jede Berührung mit der Leiche zu gewährleisten. Die Infektionsgefahr wurde somit erheblich gesteigert.
- August 2007: Uganda, Kitaka, Provinz Kamwenge, 2 Infizierte, 1 Toter
- 10. Juli 2008: Das Bernhard-Nocht-Institut für Tropenmedizin in Hamburg gibt den ersten Fall einer Einschleppung des Marburg-Virus aus Afrika nach Europa durch eine niederländische Touristin in Bakel, Niederlande, bekannt. Die 40-jährige Frau verstarb am 11. Juli 2008 in Leiden.
- 2012: Ein Ausbruch in Uganda, im Distrikt Kabale, in Ibanda im Distrikt Mbarara und im Distrikt Kampala, hält über drei Wochen an, 15 Infizierte, 4 Tote
- 2014: Uganda: Am 4. Oktober 2014 wurde der WHO ein Fall aus dem Bezirkskrankenhaus Mpigi gemeldet. Der Patient verstarb später im Krankenhaus Kampala.
- 2017: Uganda: Am 17. Oktober 2017 wurde ein Fall einer Marburg-Virus-Infektion im Distrikt Kween in Ost-Uganda der WHO gemeldet. Bis zum 14. November 2017 wurden drei Fälle, darunter ein Verdachtsfall, registriert, die alle verstarben. Der Ersterkrankte war aktiver Buschwildjäger.
- 2021: Ausbruch in Guinea: am 6. August 2021 informierte das Gesundheitsministerium Guineas die WHO über den Nachweis des Marburg-Virus bei einem Erkrankten, der seit dem 25. Juli 2021 Symptome aufwies. Die Infektion wurde nahe der Grenze zu Liberia in der Präfektur Guéckédou (Region Nzérékoré) nachgewiesen. Nachdem kein weiterer Fall aufgetreten war, erklärte die WHO den Ausbruch am 16. September 2021 für beendet.
- 2022: Ghana: Am 27. und 28. Juni 2022 verstarben in Ghana zwei Patienten aus der südlichen Ashanti Region, bei denen später eine Marburg-Virus-Infektion bestätigt wurde. Es war das erste Mal, dass das Virus in Ghana nachgewiesen wurde.

- 2023: In Äquatorialguinea wurden in der Provinz Kié-Ntem zwischen dem 7. Januar 2023 und dem 7. Februar 2023 mindestens neun durch das Marburg-Virus verursachte Todesfälle registriert.
- Tansania: Am 21. März 2023 wurde bekannt, dass in der Region Kagera bislang fünf Personen die mit dem Marburg-Virus infiziert waren gestorben sind.
Impfungen
Im April 2006 wurden Forschungsergebnisse von Forschern aus den USA und Kanada veröffentlicht, denen es gelungen ist, einen Impfstoff gegen das Marburg-Virus zu entwickeln. Im Tierversuch bei Rhesusaffen erwies sich der Impfstoff auch in der Postexpositionsprophylaxe als wirksam. Die Affen, die bei einer Infektion normalerweise nach etwa 12 Tagen verstarben, überlebten nach einer Impfung den Untersuchungszeitraum von 80 Tagen.
Therapie
Mit AVI-7288 wurde 2015 von einer amerikanischen Arbeitsgruppe erstmals ein Medikament zur Postexpositionsprophylaxe vorgestellt, das sich bei Primaten als wirksam erwies. Dabei handelt es sich um ein 23 Aminosäuren langes Phosphorodiamidate-Morpholino-Oligomer (PMO), das durch den Einsatz von Piperazine-Residuen zusätzlich positiv geladen ist (PMO plus). Dieses Oligomer bindet selektiv die mRNA-Sequenz des Nucleoproteins und unterbindet dadurch dessen Translation. Bei Makaken zeigte sich in der Placebo-kontrollierten Dosiseskalationsstudie eine dosisabhängige Wirkung bei Einsatz bis vier Tage nach Exposition mit einem Überleben bis zu 100 % bei 30 mg pro Kilogramm Körpergewicht (gegen 0 % unter Placebo). Erfahrungen bei der Verwendung am Menschen gibt es bisher nicht.
Meldepflicht
In Deutschland ist der direkte oder indirekte Nachweis des Marburgvirus namentlich meldepflichtig nach § 7 des Infektionsschutzgesetzes, soweit der Nachweis auf eine akute Infektion hinweist. Die Meldepflicht betrifft in erster Linie die Leitungen von Laboren (§ 8 IfSG).
In der Schweiz ist der positive und negative laboranalytische Befund zu einem Marburg-Virus für Laboratorien meldepflichtig und zwar nach dem Epidemiengesetz (EpG) in Verbindung mit der Epidemienverordnung und Anhang 3 der Verordnung des EDI über die Meldung von Beobachtungen übertragbarer Krankheiten des Menschen.
In Österreich ist der Verdachtsfall, Erkrankungs- und Todesfall eines virusbedingten Hämorrhagischen Fiebers meldepflichtig.
Literatur
- Ken Alibek, Stephen Handelman: Bioterror, Tod aus dem Labor. Econ, München 2001, ISBN 3-548-75089-3.
- Stephan Becker, Hans Dieter Klenk †: Das Marburg-Virus: vom Ausbruch bei den Behring-Werken bis zum Forschungsschwerpunkt an der Philipps-Universität. In: Christian Kleinschmidt (Hrsg.): Seuchenbekämpfung, Wissenschaft und Unternehmensstrategien. Die Behringwerke und die Philipps-Universität Marburg im 20. Jahrhundert (= Quellen und Forschungen zur hessischen Geschichte. Band 187). Hessische Kommission Darmstadt/ Historische Kommission Hessen, Darmstadt/ Marburg 2021, ISBN 978-3-88443-342-3, S. 235–246.
- Kristina Brauburger, Adam J. Hume, Elke Mühlberger, Judith Olejnik: Forty-five years of Marburg virus research. In: Viruses. Band 4, Nummer 10, Oktober 2012, S. 1878–1927, ISSN 1999-4915. doi:10.3390/v4101878. PMID 23202446. PMC 3497034 (freier Volltext). (Review).
- Icon Health Publications: Marburg Virus – A Medical Dictionary, Bibliography, and Annotated Research Guide to Internet Referen. Icon Health, San Diego 2004, ISBN 0-497-00706-1.
- Beate Lötfering: Das Nukleoprotein des Marburg-Virus. Untersuchungen zum Phosphorylierungsstatus. Tectum, Marburg 1998, ISBN 3-8288-0413-6.
- Christian Sänger: Untersuchungen zum Transport und zur Reifung des Marburg-Virus Oberflächenproteins GP sowie zur Ausschleusung von Nachkommenviren. Tectum, Marburg 2000, ISBN 3-8288-1045-4.
- Sven Opitz, Andrea Wiegeshoff, Malte Hagener: Das Marburg-Virus: Zur Vorgeschichte globaler Gesundheitssicherheit. In: Christian Kleinschmidt (Hrsg.): Seuchenbekämpfung, Wissenschaft und Unternehmensstrategien. Die Behringwerke und die Philipps-Universität Marburg im 20. Jahrhundert (= Quellen und Forschungen zur hessischen Geschichte. Band 187). Hessische Kommission Darmstadt/ Historische Kommission Hessen, Darmstadt/ Marburg 2021, ISBN 978-3-88443-342-3, S. 219–234.
- Richard Preston: Hot zone: tödliche Viren aus dem Regenwald; ein Tatsachen-Thriller. Droemer Knaur, München 1997, ISBN 3-426-77257-4.
- D. I. H. Simpson: Marburg and Ebola Virus Infections: A Guide for their Diagnosis, Management, and Control (= WHO Offset Publication. Nr. 36). Genf 1977 (Volltext als PDF; 952 kB).
Weblinks
- Marburg-Virus-Infektionen – Informationen des Robert Koch-Instituts
- WHO Fact sheet: Marburg haemorrhagic fever. Übersichtsseite der Weltgesundheitsorganisation (WHO, englisch)
- Marburg-Virus, auf ViralZone, Swiss Institute of Bioinformatics (SIB)
- Marburg-Virus (Memento vom 10. Juni 2010 im Internet Archive) The Universal Virus Database of the International Committee on Taxonomy of Viruses