
| Neandertaler | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
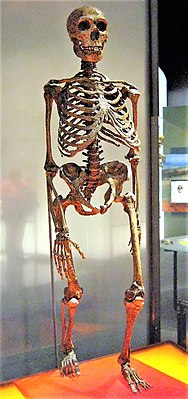
Rekonstruktion eines Neandertalerskeletts | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Pleistozän | ||||||||||||
| 230.000 (130.000) bis 30.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Homo neanderthalensis | ||||||||||||
| King, 1864 | ||||||||||||
Der Neandertaler (wissenschaftlich Homo neanderthalensis) ist ein ausgestorbener Verwandter des anatomisch modernen Menschen (Homo sapiens). Er entwickelte sich in Europa, parallel zum Homo sapiens in Afrika, aus einem gemeinsamen afrikanischen Vorfahren der Gattung Homo – dem Homo erectus – und besiedelte zeitweise große Teile Süd-, Mittel- und Osteuropas. Offensichtlich im Verlaufe der letzten Eiszeit haben die Neandertaler ihr ursprünglich ausschließlich europäisches Siedlungsgebiet bis Westasien (Türkei, Levante, Nordirak), in Teile Zentralasiens (Usbekistan, Tadschikistan) und sogar bis in das Altai-Gebiet hinein erweitert. Die DNA-Sequenzierung des Neandertaler-Erbguts ergab Hinweise auf mehrfachen Genfluss zwischen Neandertaler und Homo sapiens.
Die Neandertaler stellten Werkzeuge aus Stein und Holz her und ernährten sich – je nach klimatischen Gegebenheiten – teils von Jagdbeute, teils von Pflanzen. Sie beherrschten das Feuer, konnten sich sprachlich verständigen und waren zur Symbolbildung fähig.
Neandertaler-Funde seit der Eem-Warmzeit (vor ungefähr 130.000 Jahren) werden angesichts ihrer oft ausgeprägten anatomischen Merkmale als „klassische Neandertaler“ bezeichnet. Aufgrund zumindest vereinzelter Bestattungen ihrer Toten sowohl in Europa als auch in Westasien und des Ablegens von Toten in Höhlen ist Homo neanderthalensis neben Homo sapiens die fossil am besten überlieferte Art der Hominini. Es gibt derzeit unterschiedliche Theorien, warum die Neandertaler vor rund 40.000 Jahren ausstarben.
Namensgebung



Die Bezeichnung „Neandertaler“ geht auf das Neandertal zurück, einen zwischen den Städten Erkrath und Mettmann gelegenen Talabschnitt der Düssel. Dort, im heutigen Bundesland Nordrhein-Westfalen, wurde 1856 das Teilskelett eines Neandertalers gefunden, später Neandertal 1 benannt. Die wissenschaftliche Bezeichnung Homo neanderthalensis ist abgeleitet von lateinisch hŏmō [ˈhɔmoː] „Mensch“, das Epitheton neanderthalensis verweist – wie die volkstümlichere Bezeichnung Neandertaler – auf den Fundort. Homo neanderthalensis bedeutet somit „Mensch aus dem Neandertal“. Indirekt geht die Bezeichnung somit auf Joachim Neander zurück, nach dem das „Neandertal“ benannt wurde. Holotypus von Homo neanderthalensis ist der Fund Neandertal 1.
Die Benennung des Fossils – und damit im Ergebnis auch des Taxons – als Homo neanderthalensis erfolgte 1864 durch den irischen Geologen William King. Bereits 1863 hatte King in einem Vortrag vor der Geologischen Sektion der British Association for the Advancement of Sciences nach Erörterung der Schädelform und ihrer Abweichungen von der Schädelform des modernen Menschen den Namen „Homo Neanderthalensis King“ eingeführt. Im deutschsprachigen Raum behielt hingegen Rudolf Virchow bis zu seinem Tod im Jahr 1902 mit seiner Fehlinterpretation von 1872 die Oberhand. Virchow – der damals bedeutendste deutsche Pathologe – hielt den Fund für einen krankhaft deformierten Schädel eines modernen Menschen und verwarf die These des „Urmenschen“.
Die unterschiedlichen Schreibweisen (Epitheton „… neanderthalensis“ mit ‚th‘, Neandertaler nur mit ‚t‘) rührt daher, dass Mitte des 19. Jahrhunderts das „Neanderthal“ noch mit ‚th‘ geschrieben und diese Schreibweise seinerzeit ins Epitheton übernommen wurde. Gemäß den Internationalen Regeln für die biologische Nomenklatur werden als gültig anerkannte Artnamen nachträglich nicht mehr verändert. Die Orthographische Konferenz von 1901 legte in ihren Vorgaben für die künftige gemeinsame deutsche Orthographie aller deutschsprachigen Staaten hingegen fest, dass in heimischen Wörtern auf das bis dahin übliche ‚h‘ nach ‚t‘ grundsätzlich verzichtet werde (Tal statt Thal, Tür statt Thür). Deshalb wurde auch die bis dahin übliche volkstümliche Schreibung („Neanderthaler“) in Neandertaler verändert.
Gestützt u. a. auf eine Anregung, die der britische Paläoanthropologe Bernard G. Campbell 1973 publizierte, wurde der Neandertaler bis in die 1990er-Jahre nicht als eigene Art, sondern als Unterart von Homo sapiens angesehen und deshalb als Homo sapiens neanderthalensis bezeichnet, der anatomisch moderne Mensch als Homo sapiens sapiens. Diese Namensgebung unterstellte jedoch, dass gemäß der biologischen Nomenklatur der letzte gemeinsame Vorfahr als (archaischer) Homo sapiens zu bezeichnen wäre; tatsächlich gilt aber nach verbreiteter Sichtweise, gestützt durch neue Genanalysen (siehe Nachweis von Genfluss zum Neandertaler) der in Afrika belegte Homo erectus als letzter gemeinsame Vorfahre. Zudem hätten die Regeln der Nomenklatur zur Folge, dass – wie zum Beispiel von Günter Bräuer empfohlen – u. a. die als Homo heidelbergensis klassifizierten europäischen Vorfahren der Neandertaler ebenfalls in Homo sapiens umbenannt werden müssten. Die Einordnung des Neandertalers als Unterart von Homo sapiens gilt daher derzeit als veraltet; es gibt unter Paläoanthropologen „eine zunehmende Akzeptanz, dass die Neandertaler morphologisch unverwechselbar sind“, weswegen sich in der Fachliteratur die Bezeichnungen Homo sapiens und Homo neanderthalensis durchgesetzt haben.
Funde

Die ältesten Belege für die Anwesenheit von Individuen der Gattung Homo außerhalb von Afrika sind Fossilienfunde aus Dmanissi in Georgien, die annähernd 1,85 Millionen Jahre alt sind. Die bislang ältesten Belege für die Anwesenheit von Zuwanderern der Gattung Homo nach Europa sind 1,2 bis 1,1 Millionen Jahre alt und stammen aus der Fundstätte Sima del Elefante in der Nähe von Atapuerca (Spanien). Aus der unmittelbar benachbarten Fundstätte Gran Dolina wurden zahlreiche rund 900.000 Jahre alte Knochen geborgen. Von ihren spanischen Entdeckern werden diese Fossilien als Homo antecessor bezeichnet und in die Nähe von frühen Vorfahren der Neandertaler gestellt. Andere Forscher interpretieren diese Fossilien als Beleg für eine frühe Besiedelung der Region von Atapuerca durch eine Population von Homo erectus, die später wieder ausgestorben ist. Ob es nur eine frühe Zuwanderung nach Europa gab oder unabhängig voneinander mehrere, ist ungeklärt. In der aktuellen Fachliteratur wird überwiegend angenommen, dass Europa – lange vor der Einwanderung des Homo sapiens – von Abkömmlingen des afrikanischen Homo erectus besiedelt wurde: „Aus der europäischen Variante der Frühmenschen Homo erectus – Homo heidelbergensis genannt – gingen die Neandertaler hervor.“
Im Jahr 2006 waren insgesamt 400 Fossilfunde des Neandertalers aus Europa bekannt.
Neandertal
Mitte August 1856 entdeckten italienische Steinbrucharbeiter in einem kurz darauf dem Kalksteinabbau zum Opfer gefallenen Abschnitt des Neandertals einige Knochenfragmente. Sie wurden zunächst achtlos zum Abraum geworfen, fielen jedoch den Steinbruchbesitzern Wilhelm Beckershoff und Friedrich Wilhelm Pieper auf, die 16 größere Knochenteile bergen ließen und an Johann Carl Fuhlrott zur Untersuchung übergaben. Durch Presseberichte aufmerksam geworden, begutachtete auch der Bonner Anatom Hermann Schaaffhausen die Knochen und kam zu demselben Ergebnis wie zuvor bereits Fuhlrott: Es handele sich um eine vorzeitliche Form des modernen Menschen. Fuhlrott und Schaaffhausen präsentierten den Fund im Juni 1857 auf der Generalversammlung des Naturhistorischen Vereins der preußischen Rheinlande. Ihre Interpretation wurde jedoch vom Fachpublikum nicht geteilt. Dieser Fund, benannt Neandertal 1, ist das Typusexemplar der Art Homo neanderthalensis.
Bei Nachgrabungen an der ursprünglichen Fundstelle in den Jahren 1997 und 2000 wurden weitere 60 Knochenfragmente und Zähne entdeckt, die dem Fossil Neandertal 1 und zwei weiteren Neandertalern zugeschrieben werden konnten.
In der Nähe des Fundortes wird im Neanderthal Museum die Entwicklungsgeschichte des anatomisch modernen Menschen und seine Verwandtschaft mit dem Neandertaler nachgezeichnet.
Weitere Fundorte (Auswahl)


Der Fund aus dem Neandertal war nicht das erste entdeckte Fossil des Homo neanderthalensis. Bereits 1833 hatte der niederländische Arzt und Naturforscher Philippe-Charles Schmerling einen fossilen Kinderschädel und mehrere andere Knochen aus einer Höhle bei Engis in Belgien beschrieben, die er aufgrund von Tierfossilien und gleichfalls entdeckten Steinwerkzeugen dem „Diluvium“ (der Epoche der Sintflut) zuordnete; jedoch wurde dieser 1829 entdeckte, erste wissenschaftlich beschriebene Neandertaler-Fund (Engis 2) von den Fachkollegen als „modern“ verkannt.
Auch der 1848 im Kalksteinbruch Forbes’ Quarry in Gibraltar entdeckte, relativ gut erhaltene Schädel Gibraltar 1 wurde erst Jahrzehnte später als Jahrzehntausende alt anerkannt und zur inzwischen etablierten Art Homo neanderthalensis gestellt. Die Anerkennung des Neandertalers als eine eigenständige, von Homo sapiens abweichende Menschenform setzte sich erst endgültig durch, nachdem 1886 in einer Höhle im belgischen Spy (heute ein Ortsteil von Jemeppe-sur-Sambre) zwei fast vollständig erhaltene Neandertaler-Skelette („Homo spyensis“) gefunden worden waren.
Bis 1999 waren bereits Skelette und Skelett-Fragmente von mehr als 300 Individuen des Neandertalers bekannt. Sehr viele Fundplätze gibt es in den Karstgebieten Südfrankreichs, zum Beispiel La Chapelle-aux-Saints, Le Moustier, La Ferrassie, Pech de l’Azé, Arcy-sur-Cure und La Quina. Weitere bedeutende Fundstätten sind unter anderem die Sima de los Huesos, die Cueva de los Aviones, die Cueva Antón und die Cueva de El Sidrón in Spanien, die Tabun-Höhle und die Kebara-Höhle im Karmel-Gebirge in Israel, die Shanidar-Höhle im Irak, die Vindija-Höhle in Kroatien, die Karain-Höhle in der Türkei, die Mesmaiskaja-Höhle im russischen Teil des Kaukasus sowie die Okladnikow-Höhle im Altai-Gebirge.
Insgesamt stammt die Mehrzahl der Fossilfunde von Neandertalern – in dieser Reihenfolge – aus Frankreich, Italien und Spanien, Deutschland, Belgien und Portugal; ihr Kerngebiet war demnach Süd- und Südwesteuropa. Aus der Verteilung der bislang bekannten Fossilreste wurde abgeleitet, dass die Neandertaler erst im Verlauf der letzten Eiszeit „ihr ursprünglich ausschließlich europäisches Siedlungsgebiet bis in den Nahen Osten, in Teile Zentralasiens und sogar bis in das Altai-Gebiet hinein“ erweitert haben.
Im Jahr 2017 wurde in Science berichtet, dass im Sediment diverser gesicherter oder mutmaßlicher Aufenthaltsorte von Neandertalern Spuren ihrer mitochondrialen DNA (mtDNA) nachgewiesen werden konnten. Aus der belgischen Höhle Trou al’Wesse bei Modave waren zuvor zwar Steinwerkzeuge von Neandertalern entdeckt worden, aber keine Neandertaler-Knochen. 2021 gelang auch der Nachweis von Zellkern-DNA aus Höhlen-Sedimenten.
Fossilbericht und Alter der Funde
Die ältesten Funde im Fossilbericht, die aufgrund hinreichend vieler anatomischer Besonderheiten von der Mehrzahl der Forscher sicher als Neandertaler eingeordnet und meist als „klassische“ Neandertaler bezeichnet werden, stammen aus Grabungsschichten der Sauerstoff-Isotopenstufe MIS 5. Sie stammen aus Kroatien (Krapina) und Italien und sind etwa 130.000 bzw. 120.000 Jahre alt. Der namensgebende Fund aus dem Neandertal wurde auf ein Alter von 42.000 Jahren datiert.

Die Abgrenzung der zu den Neandertalern gestellten Knochenfunde von den früher als Vor-Neandertaler („Ante-Neandertaler“, „Prä-Neandertaler“, „Proto-Neandertaler“) und heute meist als Homo heidelbergensis bezeichneten älteren Funden ist schwierig, da die Neandertaler unmittelbar und allmählich aus der Chronospezies Homo heidelbergensis hervorgingen. Daher werden in den Fachpublikationen unterschiedliche Datierungen ausgewiesen. Häufig wird die Existenz der Neandertaler als eigenes Taxon von Fossilien hergeleitet, die zwischen 200.000 und 160.000 Jahre alt sind; vereinzelt wurden aber auch 300.000 Jahre und sogar 500.000 Jahre alte Fossilien den Neandertalern zugeschrieben.
Nicht sicher datierbar ist bisher auch der Zeitpunkt, an dem die Neandertaler ausstarben. Der verbreiteten Ansicht, dass die Neandertaler besonders an die Kälteperioden (Stadiale) der vor rund 115.000 Jahren einsetzenden letzten Kaltzeit angepasst waren, scheint zu widersprechen, dass sie offenbar während eines Interstadials, des Interstadials 5, ausstarben. Dies war lange vor dem glazialen Maximum der letzten Kaltzeit, das vor rund 25.000 Jahren begann und seinen Höhepunkt vor rund 20.000 Jahren erreichte. Auch die Verteilung der Knochenfunde wurde dahingehend interpretiert, dass die Neandertaler nur „unter günstigen klimatischen Bedingungen und Umweltverhältnissen“ ihr im Südwesten und Süden Europas gelegenes Kerngebiet verlassen haben, „um in Gebiete vorzudringen, in denen sie sich nur zeitweilig, bis zur Verschlechterung der dortigen klimatischen Bedingungen bzw. Umweltverhältnisse, aufhielten.“ Einer 2011 veröffentlichten Studie zufolge markieren die Fossilien aus der kaukasischen Mesmaiskaja-Höhle (39.700 ± 1.100 cal BP) die jüngsten Neandertalerfunde mit unzweifelhafter Datierung. Auch die Datierung von Funden aus der Iberischen Halbinsel, die mit jünger als 45.000 Jahre bestimmt wurden, gilt als zweifelhaft.
Umstritten ist ferner die Zuverlässigkeit der Altersbestimmung von anderen, jünger datierten Funden; dies betrifft insbesondere Funde der Höhlen von Arcy-sur-Cure (34.000 Jahre vor heute = BP), aus der Cueva del Boquete de Zafarraya (32.000 BP) und aus der Gorham-Höhle (28.000 BP). Diese Fundorte liegen zudem alle deutlich südlicher und sprechen daher eher für eine Kälteflucht. Datierungen von Neandertaler-Fossilien jünger als 34.000 BP (14C-Jahren) werden entweder aus methodischen Gründen oder wegen der Überlieferung aus einem unklaren Schichtzusammenhang angezweifelt. Möglicherweise wurde das Alter der in Südspanien gefundenen Fossilien aufgrund von Verunreinigungen bei der Probenentnahme um rund 10.000 Jahre zu jung eingestuft;Thomas Higham, ein britischer Experte für Radiokarbon-Datierungen, geht aufgrund diverser von ihm vorgenommener Altersbestimmungen davon aus, dass die Neandertaler spätestens vor 39.000 Jahren (cal BP) in Europa ausgestorben waren. Umstritten ist auch die Zuordnung von Moustérien-artigen Steinwerkzeugen zum späten Neandertaler, die auf 65° 01′ N (also fast am Polarkreis) im nördlichen Ural in der Fundstelle Byzovaya entdeckt und auf ein Alter von 34.000 bis 31.000 BP datiert wurden.
Vor etwa 40.000 Jahren drang Homo sapiens aus Afrika über den Nahen Osten nach Norden vor und besetzte in der Folge den bisherigen Lebensraum der Neandertaler. Die Kultur des Châtelperronien gilt als Beleg für die kulturelle Beeinflussung der Neandertaler durch die anatomisch modernen Menschen (Homo sapiens) des Jungpaläolithikums, die sogenannten Cro-Magnon-Menschen.
Körperbau
Dank der mehr als 300 Skelettfunde ist der Neandertaler die am besten untersuchte fossile Art der Gattung Homo. Gleichwohl wies Ian Tattersall darauf hin, dass es bis Ende der 1970er-Jahre nur „eine vordergründige Definition“ dieses Taxons gab; gefehlt habe jedoch eine Zusammenstellung jener Merkmale, die Homo neanderthalensis von allen anderen Arten der Gattung Homo unterscheiden. Diese habe erst Albert Santa Luca 1978 vorgelegt und dabei vier einzigartige Merkmale des Neandertaler-Schädels herausgestellt:
„Eines war der Torus occipitalis (Hinterhauptswulst), eine knöcherne Leiste, die quer über das Hinterhauptsbein am Hinterkopf verläuft. Über diesem Wulst liegt eine ovale Vertiefung (Fossa suprainiaca), ein weiteres ausschließliches Neandertaler-Merkmal. Weiter vorn an der Schädelbasis findet sich das dritte Merkmal, ein ausgeprägter occipito-mastoidaler Kamm (heute oft als Juxtamastoid-Kamm bezeichnet), dieser liegt im Mastoid-Fortsatz. Der Mastoid-Knochen ist eine Knochenstruktur, die (bei Neandertalern im Vergleich zum anatomisch modernen Menschen klein) hinter und unter dem Ohrkanal vorspringt. Schließlich besitzen Neandertaler oben auf dem Mastoid-Fortsatz eine deutliche, gerundete Erhöhung, die Tuberositas mastoidalis. Diese schräg nach hinten und oben verlaufende Erhöhung ist bei anderen Menschenformen anders entwickelt oder fehlt.“
Später fand man weitere Neandertaler-typische Merkmale, beispielsweise spezielle Strukturen der Nasenhöhle und die Lage der Bogengänge des Innenohrs. Analysen zweier gut erhaltener Skelette von Neandertaler-Neugeborenen zeigten, dass die – im Vergleich mit dem anatomisch modernen Menschen – robusten Knochen der Neandertaler bereits vor der Geburt angelegt waren.
Fußspuren der Neandertaler sind insbesondere aus einem Dünengebiet von Le Rozel in der Normandie (Frankreich, rund 70.000 Jahre alt) bekannt; dort hat eine Gruppe von 10 bis 13 vorwiegend sehr junger und jugendlicher Neandertaler mindestens 257 Abdrücke hinterlassen. 87 Abdrücke, die 2020 im Nationalpark Coto de Doñana im Südwesten von Spanien entdeckt wurden (106.000 ± 19.000 Jahre alt), stammen ebenfalls von einer Gruppe zumeist junger Neandertaler. Ein einzelner Fußabdruck ist ferner zum Beispiel aus der Vârtop-Höhle im Bihor-Gebirge (Rumänien mindestens 62.000, maximal 97.000 Jahre alt) erhalten geblieben.
Schädelknochen


Das Gehirnvolumen der Neandertaler betrug rund 1200 bis 1750 Kubikzentimeter (im Mittel rund 1400 cm³), was im Durchschnitt etwas größer ist als beim heutigen Menschen und als eine Folge ihres insgesamt kräftigeren Körperbaus gedeutet wird. Diese Variationsbreite hatte zur Folge, dass auch andere Merkmale des Schädels eine erhebliche Variationsbreite aufweisen. Gleichwohl gibt es zahlreiche Merkmale, die sich von jenen des anatomisch modernen Menschen unterscheiden und die sich zudem nicht erst nachgeburtlich herausbildeten, sondern bereits vorgeburtlich angelegt waren; dies konnte am Schädel des Neandertaler-Babys aus der Mesmaiskaja-Höhle (Kaukasus) belegt werden.
Der Schädel ist von vorn nach hinten länglich geformt und mit seiner niedrigen Stirn auch viel flacher als bei heutigen Menschen, zudem hat seine längliche Form zur Folge, dass er weit nach hinten ragt und dort einen charakteristischen Vorsprung bildet. Aufgrund seiner beiden stark vorspringenden Überaugenwülste wirkt die Schädelform archaischer als die der meisten heute lebenden Menschen. Die größte Schädelbreite liegt auf Höhe der unteren Schädelbasis (beim anatomisch modernen Menschen: über den Ohren). Dadurch und durch den verhältnismäßig niedrigen, breiten Hirnschädel erscheint der Umriss in der Ansicht von hinten als halbkreisförmig (beim anatomisch modernen Menschen: abgerundet trapezförmig). Am Gesichtsschädel ist außerdem die große und breite Nasenöffnung auffällig.
Ein besonders trennscharfes Merkmal zwischen Neandertaler und Homo sapiens bildet die Lage der Bogengänge des Innenohrs im Felsenbein der Schädelbasis. Der halbkreisförmige hintere Bogengang (ein Anteil des Gleichgewichtsorgans für Drehbeschleunigungen) liegt beim Neandertaler tiefer als bei sämtlichen anderen Arten der Gattung Homo. Der Unterschied zwischen Neandertalern und Homo sapiens ist bei diesem Merkmal etwa so groß wie der zwischen Homo sapiens und Schimpansen.

Die Stirn ist flach und fliehend, während sie beim europäischen Homo sapiens meist steil ist. Die Region über den Augen zeigt typischerweise einen deutlichen Überaugenwulst (Torus supraorbitalis). Jedoch sind die Überaugenwülste nicht bei allen Individuen stark ausgeprägt, kamen zudem auch beim frühen Homo sapiens vor und sind daher nicht immer ein verlässliches Kriterium zum Unterscheiden von Neandertaler und Homo sapiens. Diese Knochenverdickung wird als stabilisierende Anpassung gedeutet, denn der Schädel war – durch den kräftigen Kauapparat – starken statischen Belastungen ausgesetzt. Das Merkmal trat bereits bei den gemeinsamen Vorfahren von Neandertalern und anatomisch modernen Menschen auf und ist auch bei den Menschenaffen zu beobachten.
Die Nasenöffnung ist groß und relativ breit, die Nasenwurzel sehr kräftig und ebenfalls breit. Der Nasenboden mündet, im Gegensatz zum modernen Europäer, abgerundet in die Gesichtsebene. Diese Merkmale deuten auf eine große, fleischige Nase hin, was – wie einige weitere, innere Nasenmerkmale – von einigen Forschern als Anpassung an die eiszeitlichen Kälteperioden gedeutet wird: Eine große Nase wärme die Atemluft vor, bevor diese die Lungen erreicht, und unterstütze so das Aufrechterhalten der Körper-Kerntemperatur. Zudem war die Riechschleimhaut weiter vorn in der Nase angeordnet als bei Homo sapiens: „Die damit verbesserte Aufnahme von Gerüchen könnte allgemein ein Vorteil bei der Ortung von Nahrung und speziell bei der Jagd auf Tiere gewesen sein.“ Argumentiert wurde aber auch, dass die größeren Nasen sowie die geräumigeren Kieferhöhlen und Stirnhöhlen der Neandertaler weniger eine Anpassung an Kältezeiten waren, sondern primär eine Folge ihres insgesamt etwas breiteren Gesichts.
Bezahnung


Anhand der Abnutzung der Zähne wurde geschlossen, dass die Krapina-Neandertaler nicht älter als 30 Jahre wurden; nur unwesentlich höhere Lebensspannen wurden auch für die Fossilien von Homo heidelbergensis aus der Sima de los Huesos in Spanien bekannt.
Oberkiefer- und Unterkieferknochen sind höher und auch länger als beim anatomisch modernen Menschen; auch die Schneidezähne der Neandertaler sind größer, die Backenzähne jedoch schmaler als bei Homo sapiens. Bedingt durch die kräftigeren und größeren Kieferknochen wirken Neandertalerschädel prognath, d. h. die untere Gesichtshälfte ragt deutlich hervor. Die aufsteigenden Unterkieferäste sind breiter, der Winkel zwischen Unterkieferästen und -körper steiler. Ein auffälliges Unterscheidungsmerkmal zum anatomisch modernen Menschen ist bei den meisten Neandertalerschädeln das Fehlen eines deutlich vorspringenden Kinns.
Die Anzahl der Zähne sowie die Kronenformen stimmen mit denen des Homo sapiens überein, jedoch sind die oberen Schneidezähne schaufelförmig gekrümmt. Die Backenzähne haben häufig einen Höcker in ihrer Mitte, der beim anatomisch modernen Menschen nicht vorkommt. Die hinteren Backenzähne sind zuweilen – nicht immer – durch Taurodontie gekennzeichnet, d. h. die Wurzeln trennen sich erst kurz vor den Spitzen in Äste auf. Besondere diagnostische Merkmale sind außerdem an den unteren vierten Prämolaren, den ersten Molaren und zweiten Milchmolaren festzustellen, was inzwischen zu umfangreichen vergleichenden Studien an spätmittelpaläolithischen und frühjungpaläolithischen Zahnfunden zur Unterscheidung von Neandertalern und anatomisch modernen Menschen geführt hat. Außerdem ist die sogenannte retromolare Lücke („Neandertaler-Lücke“, nicht zu verwechseln mit dem Diastema) typisch, die regelmäßig zwischen dem letzten Molaren (Backenzahn) und dem Unterkieferast auftritt.
Eine Hypothese geht davon aus, dass die Form des Schädels nicht nur passiv durch das in ihm heranwachsende Gehirn geformt wurde, sondern später auch durch die starke Beanspruchung der Schneidezähne zustande kam. Diese wurden demnach nicht nur zur Nahrungsaufnahme, sondern auch als „Werkzeug“ und als eine Art „dritte Hand“, benutzt. Die sogenannte Teeth-as-tool-Hypothese von F. H. Smith besagt, dass die Zähne als Schraubstock und Zange eingesetzt wurden. Jedoch ist diese Technik kein Alleinstellungsmerkmal der Neandertaler, sondern sowohl pathologisch als auch ethnografisch beim modernen Menschen belegt. Abriebspuren an den Zähnen deuten darauf hin, dass die Neandertaler – wie schon Homo heidelbergensis – überwiegend Rechtshänder waren.
Rumpf, Arme und Beine
Viele Neandertaler-Funde stammen aus Bestattungen, weswegen sämtliche Bereiche ihres Körpers mehrfach und in gutem Zustand überliefert wurden. Die typischen europäischen – die so genannten klassischen – Neandertaler-Skelette „sehen mehr oder weniger genauso aus wie die Skelette heutiger Menschen. Unterschiede bestehen vor allem in den Proportionen. Neandertaler haben ein viel breiteres, robuster gebautes Becken, und auch die Beinknochen sind kräftiger als bei heutigen Menschen. Dagegen waren die Arme vergleichsweise zierlich gebaut.“ Aus den Muskelansatzmarken der Hände wurde abgeleitet, dass Neandertaler bei ihren händisch ausgeführten Tätigkeiten primär Präzisionsgriffe einsetzten.
Die Knochenfunde lassen auf Körpergrößen von ca. 1,60 m schließen; die Neandertaler waren demnach etwas kleiner als die frühen anatomisch modernen Menschen, für die eine Körpergröße von ca. 1,77 m rekonstruiert wurde. Ihr Körpergewicht entsprach hingegen ungefähr dem der heute lebenden Europäer: Dem so genannten Alten Mann von La Chapelle, einem 1908 im französischen La Chapelle-aux-Saints (Département Corrèze) gefundenen Schädel mit zugehörigem Unterkiefer und zahlreichen weiteren Körperknochen, wird ein Körpergewicht von 60 bis 80 kg zugeschrieben; dem 1848 in Gibraltar im Forbes’ Quarry entdeckten weiblichen Schädel Gibraltar 1 wird ein Körpergewicht von 50 bis 70 kg zugeschrieben. Die Körpergröße der Neandertalerinnen betrug ungefähr 95 Prozent der durchschnittlichen Größe von Neandertaler-Männern und entspricht somit den Verhältnissen beim modernen Menschen. Der Beckenkanal der Neandertalerinnen war ähnlich eng gebaut wie der von anatomisch modernen Frauen.
Angesichts der Tatsache, dass die Neandertaler während einer Eiszeit lebten, wurden solche Unterschiede als Anpassung an das kalte Klima in Europa gedeutet. Funde aus wärmeren Gegenden (zum Beispiel dem Nahen Osten) weisen auf größere und schlankere Individuen hin. Da zwischen Brustkorb und Hüfte der Neandertaler nur ein kurzer Zwischenraum war und die Brusthöhle durch eine vom anatomisch modernen Menschen abweichende Biegung der Rippen größer war als bei Homo sapiens, wirkte ihr Rumpf kompakter, stämmiger – „fassförmiger“ – als der Rumpf heutiger Europäer; dies gilt auch als der Hauptgrund für die im Vergleich mit den heute lebenden Menschen im Durchschnitt geringere Körpergröße der Neandertaler.

Als Anpassung des Körperbaus an ein relativ kaltes Klima werden auch die Abweichungen bestimmter Merkmale der Beine vom Jetztmenschen gedeutet; Friedemann Schrenk verdeutlichte dies am Beispiel von Afrikanern, Lappen und Neandertalern:
„Während bei den ‚Lappen‘ der Unterschenkel in der Länge 79 Prozent des Oberschenkels entspricht, liegt dieser Wert bei den Afrikanern bei 86 Prozent; diese haben also weitaus längere Unterbeine. Die Unterschenkel der Neandertaler entsprachen in der Länge nur 71 Prozent des Oberschenkels, also hatten die Neandertaler deutlich kürzere Beine als heutige Menschen aus Lappland.“
Neben diesen gegenüber dem Homo sapiens abweichenden Längenverhältnissen waren die Knochen der unteren Extremitäten bei den Neandertalern auch weitaus größeren Belastungen gewachsen:
„[Oberschenkelknochen und Schienbein] lassen auf eine Verdoppelung der Biege- und Torsionsbelastbarkeit im Vergleich zur unteren Extremität moderner Menschen schließen. Die Morphologie des Kniebereichs weist auf beachtliche Kräfte und Belastungsfähigkeit hin. Schließlich war der Fuß aufgrund vergrößerter Gelenke und einer verstärkten Großzehe extrem beanspruchbar.“
Allerdings wurde aus der Länge seiner Achillessehne abgeleitet, dass der Neandertaler ein weniger guter Ausdauerläufer war und auch beim Kurzstreckenlauf mehr Energie verbrauchte als der moderne Mensch.
Aus den erhaltenen Muskelmarken (den Ansatzstellen der Muskeln am Knochen) konnte abgeleitet werden, dass die Neandertaler im Vergleich zum Jetztmenschen eine ungewöhnlich starke Brust- und Rückenmuskulatur hatten, so dass die Arme „auch einen überaus starken Kraftgriff“ erlaubten; die Handknochen lassen zusätzlich auf einen „Präzisionsgriff“ schließen. Von diesen Muskelmarken und dem Gewicht der Knochenfunde – auch die Rippen und der Beckengürtel waren massiver geformt als beim modernen Menschen – konnte auf das Körpergewicht zurückgeschlossen werden, das mit 50 bis 80 kg im Verhältnis zur Körpergröße und im Vergleich zum heutigen Menschen relativ hoch ist.
Das früher häufig dargestellte Bild vom schwerfälligen Primitiven, der kaum aufrecht gehen kann, ist längst überholt, denn die Körpermaße der Neandertaler liegen – trotz aller Abweichungen – noch innerhalb der Variationsbreite heutiger Menschen.
Anatomische Befunde zu Entwicklung und Sozialverhalten

Forscher des Max-Planck-Instituts für evolutionäre Anthropologie wiesen anhand von Abdrücken des Gehirns auf der Innenseite der Schädelknochen nach, dass sich das Wachstumsmuster des Gehirns der Neandertaler im ersten Lebensjahr – einer kritischen Phase für die kognitive Entwicklung – erheblich von dem des anatomisch modernen Menschen unterschied. Demnach hatte die leicht unterschiedliche Form des Gehirns (bei modernen Menschen kugelig, bei Neandertalern länglich) vermutlich Auswirkungen auf die kognitiven Fähigkeiten. So entwickelten sich beim Neandertalerkind offenbar Bereiche des Parietallappens und der Kleinhirnregion weniger stark als beim jungen Homo sapiens, aber ähnlich wie beim Schimpansen. Wenn diese Regionen beim modernen Menschen verletzt werden oder infolge von Entwicklungsstörungen verkleinert sind, kann dies zu Einschränkungen beim Sprechen und beim Sozialverhalten führen.
In einem Laborexperiment integrierten Zellbiologen die für Neandertaler rekonstruierte Variante des Gens NOVA1 in Stammzellen eines heutigen Menschen; von diesem Gen ist bekannt, dass es die Funktionsweise des Gehirns in frühen Phasen der Entwicklung reguliert. Daraufhin wuchs ein „Organoid“ heran, dessen Merkmale sich deutlich von einem unveränderten Organoid unterschieden: Es hatte eine andere Form, und „die Art, wie sich die Zellen ausbreiten, wie sich die Synapsen als Verbindungsstellen der Neuronen bilden und auch, aus welchen Proteinen die Synapsen aufgebaut sind“, war ebenfalls unterschiedlich. Daraus schlossen die Forscher, dass das Gehirn der Neandertaler sich merklich von dem des Homo sapiens unterschieden hat.
Mit Hilfe von Synchrotronstrahlung wurde im Jahr 2010 rekonstruiert, innerhalb welcher Zeitspanne sich die Zähne der Neandertalerkinder entwickelten; dies gilt beim anatomisch modernen Menschen als Maßstab für die generelle Entwicklungsgeschwindigkeit eines Kindes. Demnach war die Entwicklungsgeschwindigkeit der jungen Neandertaler wesentlich rascher – und die Phase der Kindheit somit kürzer – als beim Menschen. Allerdings verlief die Entwicklung des Gehirns in früher Kindheit vermutlich ähnlich wie beim anatomisch modernen Menschen, und auch die Vergrößerung der Knochen unterhalb des Kopfes ähnelte einer 2017 publizierten Studie zufolge dem Verlauf beim Menschen; dies wurde als Hinweis auf eine möglicherweise ähnlich lange Kindheit wie beim Menschen interpretiert.
Die rechten Oberarmknochen und die rechtsseitigen Muskelansätze der Oberarme von Neandertalern waren in der Regel kräftiger ausgebildet als die linken. Dies wird häufig auf den regelmäßigen Gebrauch von Speeren zurückgeführt; eine 2012 publizierte Studie legte jedoch nahe, dass diese ausgeprägte Asymmetrie vor allem eine Folge des häufigen Bearbeitens von Oberflächen (Glätten des Bodens, von Fellen) sein könnte.
Zahlreiche Skelette älterer Neandertaler weisen verheilte Knochenbrüche und Hinweise auf stark zurückgebildete Muskeln als Folge von Verletzungen auf, die sie erheblich schwächten. Dies wurde dahingehend interpretiert, dass sie die Folgen dieser Verletzungen nur überleben konnten, weil sie von Sippenmitgliedern unterstützt wurden.
In den Höhlen von Goyet in Gesves (Belgien) wurden – wie zuvor in Ausgrabungsstätten in Frankreich und Spanien – Hinweise auf Kannibalismus gefunden.
Lebenserwartung
Das Alter bei Eintritt des Todes kann zwar für einzelne Knochenfunde recht genau rekonstruiert werden. Ein verlässlicher Mittelwert für die Lebenserwartung der gesamten Neandertaler-Population kann hieraus aber nicht berechnet werden. Dennoch gibt es Anhaltspunkte für die Lebenserwartung. Friedemann Schrenk zufolge ergab eine Untersuchung „von insgesamt 220 Skeletten aus dem gesamten Verbreitungsgebiet der Neandertaler aus einem Zeitraum von vor 100.000 und 35.000 Jahren, […] daß 80 Prozent aller Neandertaler vor dem 40. Lebensjahr starben“. Obwohl die meisten von ihnen sogar schon im Alter zwischen 20 und 30 Jahren starben, wurden immerhin 20 Prozent (also jeder fünfte Neandertaler) älter als 40 Jahre, „was eine beachtliche Leistung darstellt“. Als gesichert gilt beispielsweise, dass der sogenannte Alte Mann von La Chapelle ungefähr 40 bis 45 Jahre alt war, als er starb. Gleichwohl deuten diese Befunde darauf hin, dass nur wenige Personen noch das Aufwachsen ihrer Enkel miterlebten. (Vergl. auch Großmutter-Hypothese)
Ernährung
Die Zähne von heute lebenden Menschen und Makaken weisen ein Merkmal auf, das streng mit dem Zeitpunkt des Abstillens korreliert: das Verhältnis von Barium zu Kalzium im Zahnschmelz. Eine Analyse dieses Verhältnisses in einem Neandertaler-Zahn ergab im Jahr 2013, dass dieser Neandertaler im Alter von rund 14 bis 15 Monaten abgestillt worden war. Die Untersuchung von zwei Neandertaler-Zähnen aus der französischen Fundstelle Payre (Gemeinde Rompon, Département Ardèche) ergab 2018 hingegen ein Alter von 2 ½ Jahren für den Zeitpunkt des Abstillens; 1997 war das Alter bei Entwöhnung auf drei Jahre geschätzt worden. Eine 2020 publizierte Studie an drei, rund 70.000 bis 50.000 Jahre alten Neandertaler-Zähnen aus Italien, ergab hingegen ein Alter von nur 5 bis 6 Monaten für das Abstillen.

Gesichert ist, dass die Neandertaler an ihren Wohnplätzen regelmäßig Feuer entfachten; die ältesten, als gesichert geltenden Feuerstellen in Europa stammen bereits von Homo heidelbergensis und sind rund 400.000 Jahre alt. Besonders aufschlussreich waren Ascheablagerungen von einer Fülle von Feuerstellen, die in der Kebara-Höhle entdeckt wurden: „Jede Siedlungsphase hinterließ in der Höhle eine Abfallschicht; in der Zeit zwischen den Wohnphasen wehte Staub hinein und Felsmaterial fiel von der Decke. In Kebara haben sich meterdicke Sedimente angesammelt, in denen man genau in der Zentralfläche, wo die Feuerstellen lagen, aufeinanderfolgende Begehungshorizonte unterscheiden konnte.“ Ähnliche Funde wurden in Spanien im Abric Romaní freigelegt, einem Felsvorsprung (Abri), der – mit Unterbrechungen – mehr als 20.000 Jahre lang bewohnt wurde. Fundstätten von Jagdplätzen in Frankreich, im Kaukasus und bei Wallertheim im Rheinland sowie in der Jagdstation Buhlen belegen, „dass Neandertaler spezialisierte Jäger waren, die Bisons oder Mammuts auf ihren Wegen in Winterweidegebiete immer wieder an denselben Stellen auflauerten und erlegten. In Salzgitter-Lebenstedt fanden sich zusammen mit tausenden Steinwerkzeugen Knochenreste von 86 erjagten Rentieren, ein eindeutiges Zeugnis für die ausgezeichneten Jagdfähigkeiten der Neandertaler.“
Der Anteil von verzehrtem Fleisch an der Nahrung war jedoch offenbar regional und in unterschiedlichen Epochen nicht gleichförmig. Isotopenmessungen von Kollagen in Neandertalerknochen aus der Vindija-Höhle in Kroatien deuten beispielsweise darauf hin, dass Fleisch die hauptsächliche Quelle für Protein war; Isotopenmessungen bei mehreren Neandertaler-Funden aus Frankreich wurden sogar dahingehend interpretiert, diese Neandertaler seien „Spitzenprädatoren“ gewesen. Derartige Befunde führten unter anderem zur Vermutung, das Aussterben der Neandertaler könne durch eine im Vergleich zu Homo sapiens weniger flexible Ernährung mitverursacht worden sein. Im Jahr 2010 wurde diese Hypothese jedoch abgeschwächt, als ein internationales Forscherteam vom Center for Advanced Study of Hominid Paleobiology der George Washington University im Zahnstein von Neandertaler-Zähnen aus Belgien und dem Irak zahlreiche pflanzliche Mikrofossilien nachweisen konnte. Demnach wurden unter anderem Datteln, Hülsenfrüchte und Grassamen verzehrt. Ferner wurde festgestellt, dass die im Zahnstein eingelagerte Stärke von nordspanischen Neandertalern Merkmale einer Veränderung durch Erhitzung aufweist; deren pflanzliche Kost war folglich durch Kochen verdaulicher gemacht worden, und sie bestand zumindest teilweise aus Arten, die als Heil- oder Gewürzpflanzen interpretiert werden können. In Shanidar (Irak) wurden Reste von erhitzten Pflanzen beschrieben, die als Beleg für das Herstellen von Mahlzeiten vor rund 70.000 Jahren interpretiert wurden.
Auch anhand von Abriebspuren auf der Zahnoberfläche von Neandertalern aus unterschiedlichen Epochen (Kältezeiten und Warmzeiten) wurde belegt, dass sie je nach klimatischen und damit zugleich ökologischen Gegebenheiten ihre Nahrungsaufnahme an das jeweils vorhandene Pflanzen-Angebot anpassten. Unabhängig von diesen Befunden konnte auch aus rund 50.000 Jahre altem Kot von spanischen Neandertalern – anhand von erhaltenem 5β-stigmasterin – rekonstruiert werden, dass neben häufigem Fleischkonsum auch ein erheblicher Anteil an pflanzlicher Kost verzehrt wurde. Eine Isotopenuntersuchung der Skelettfunde von Spy (heute Ortsteil von Jemeppe-sur-Sambre in Belgien) ergab, dass rund 20 Prozent der Protein-Aufnahme pflanzlicher Herkunft war, was zugleich bedeutet, dass diese Population einen höheren Fleischanteil in der Nahrung aufwies als die Populationen in Spanien. Gleichwohl belegt die Analyse eines rund 140.000 Jahre alten Zahns aus der Provinz Huesca im Nordosten Spaniens, dass es auch dort in jener Epoche einen lebenslang hohen Fleischkonsum gab.
Eine Arbeitsgruppe des Forschungsinstituts Senckenberg analysierte im Jahr 2011 die Abnutzungsspuren von 73 Backenzähnen aus dem Oberkiefer von Neandertalern und modernen Menschen: Durch das Zerkleinern der Nahrung verändert sich die Zahnoberfläche in Abhängigkeit vom Nahrungstyp. Die Untersuchungsergebnisse „zeigen eindeutig, dass die Nahrung bei beiden Vertretern der Gattung Homo insgesamt vielseitig ausfiel.“ Zudem wurde nachgewiesen, dass die Zusammensetzung der Nahrung „jeweils von den öko-geografischen Gegebenheiten abhing.“ Auch dieser Studie zufolge war der Fleischanteil in der Nahrung bei den in Nordeuropa lebenden Neandertalern wesentlich höher als bei den in Südeuropa lebenden Neandertalern. Die Nahrung der Neandertaler war demnach ähnlich variabel und abwechslungsreich wie die des frühen europäischen Homo sapiens.
Die bislang ältesten Belege für den Verzehr von Schnecken und Muscheln stammen aus der Bajondillo-Höhle (Torremolinos, Spanien); sie wurden der Sauerstoff-Isotopenstufe MIS 6 zugeordnet und auf ein Alter von 150.000 Jahren datiert. Belege für die Anpassung der Neandertaler an öko-geografische Gegebenheiten wurden auch in der küstennahen Vanguard-Höhle und der ebenfalls küstennahen Gorham-Höhle in Gibraltar entdeckt, gemeinsam mit Steinwerkzeugen aus dem Moustérien: Schalen von adriatischen Miesmuscheln (Mytilus galloprovincialis) sowie Knochen von Robben, Delfinen und Fischen zeugen dort für einen vieltausendjährigen Verzehr von Meerestieren. Auch in der Serra da Arrábida in Portugal wurden rund 100.000 Jahre alte Belege für den Verzehr von Meerestieren entdeckt.Gehörgangsexostosen bei einigen untersuchten Schädeln wurden als unabhängige Bestätigung dafür gedeutet, dass Neandertaler Nahrung auch unter Wasser gewinnen konnten. Aus der Gorham-Höhle stammen auch die ältesten Belege für das Verzehren von Tauben durch Neandertaler; die erhalten gebliebenen Knochen dieser Vögel weisen sowohl Schnitt- als auch Brandspuren auf.
Kultur

Übersicht
Lebensraum und Kultur von Homo neanderthalensis erstreckten sich – insbesondere in der Phase als „klassischer Neandertaler“ seit der Eem-Warmzeit vor ca. 125.000 Jahren – über weite Teile Europas bis zur Levante im Nahen Osten und über die Krim-Halbinsel hinaus bis an den Rand Sibiriens, wobei Sibirien archäologischen Befunden zufolge vermutlich in zwei Wellen besiedelt wurde. Diese frühen Europäer lebten in arbeitsteiligen Gruppen. Die Neandertaler werden insbesondere mit dem Kulturraum des Moustérien (vor 125.000 bis 40.000 Jahren) mit u. a. Micoquien- und Levalloistechnik der Steinbearbeitung in Verbindung gebracht – bis in die Zeit ihres Verschwindens im frühen Aurignacien, in dem bereits der anatomisch moderne Homo sapiens (Cro-Magnon-Mensch) in Europa zugewandert war. Die Horden siedelten z. T. weit verstreut, und es kann nicht von einer einheitlichen Lebensweise in diesem riesigen Gebiet ausgegangen werden. Auch ein einheitliches Erscheinungsbild der Individuen ist unwahrscheinlich, wenngleich vereinzelt genetische Spuren für rotes Haar und hellere Pigmentierung nachgewiesen wurden.
Regional unterschiedliche Gegebenheiten bestimmten den Lebensalltag der Neandertaler: Klima, Gelände und Jahreszeiten, Trinkwasservorkommen und Vorhandensein des jagdbaren Wildes und anderer Nahrungsmittel, insbesondere Plätze, an denen Rohstoffe für Steinwerkzeuge vorkamen. Manche Gruppen hielten sich vorzugsweise in Höhlen und Grotten oder unter Abris (Felsüberhängen) auf – z. B. in der Dordogne (Le Moustier, La Ferrassie) aber auch in der Kleinen Feldhofer Grotte im Neandertal. Andere lebten in der Ebene oder in Waldungen und bauten sich Unterschlupfe aus Fellen oder Strauchwerk und Ästen. Auch gab es Behausungen, die mit Mammutknochen und Stoßzähnen abgestützt wurden, z. B. im Netzetal (Hessen). In Rheindahlen bei Mönchengladbach wurden flache Gruben mit runden Stützlöchern und Feuerstellen gefunden, auf einem Vorplatz Steinartefakte aus mehreren Zonen: grob zerlegte Steinknollen sowie feine Kantenbearbeitung durch Retuschierung. Diese Funde stammen aus der Eem-Warmzeit. Auch in der Ukraine gab es Freilandstationen mit Belegen für Feuerstellen.
In Frankreich wurden an Faustkeilen und Abschlägen Spuren von Pyrit gefunden. Wissenschaftler, die jene Werkzeuge nachgebildet haben und mit diesen Feuer erzeugten (was ebenfalls zu Pyrit-Spuren am Werkzeug führte), schlossen daraus, dass Äxte und andere Steinwerkzeuge von Neandertalern teilweise multifunktionell verwendet wurden.
Im Nahen Osten zeigten Neandertaler unterschiedliches Wanderverhalten: Zum einen gab es Rundwanderstrategien von Ort zu Ort, zum anderen sternförmige Wanderungen vom Basislager zu peripheren Plätzen mit Rohstoffvorkommen. Im Mittelpaläolithikum suchten die Neandertaler gezielt größere Lagerstätten von Feuerstein und Quarzit auf, an einigen Orten über zehntausend Jahre.
Steinwerkzeuge und Waffen, die heute bestimmten Kulturen oder Bearbeitungstechniken zugeordnet werden (Faustkeile, Abschläge, Schaber, Spitzen) wurden nicht immer von allen Neandertalergruppen benutzt und nicht immer im gleichen Zeitraum. Manche kamen überwiegend in einer bestimmten Region vor. Gegen Ende ihrer Existenz wurden die Techniken der Neandertaler möglicherweise durch Werkzeuge und Schmuckobjekte eingewanderter Cro-Magnon-Menschen beeinflusst.
Analysen des Erbguts von zwei ca. 120.000 Jahre alten Neandertalern aus Deutschland und Belgien ergaben, dass die letzten Neandertaler, die vor rund 40.000 Jahren in Europa lebten, zumindest teilweise von diesen ca. 80.000 Jahre älteren europäischen Neandertalern abstammten. Zugleich ergaben die Analysen, dass die beiden 120.000 Jahre alten Individuen weniger eng mit den zur selben Zeit in Sibirien lebenden Neandertalern verwandt waren, was bedeutet, dass die Neandertaler-Populationen in Europa und Sibirien bereits vor 120.000 Jahren kaum noch Kontakt untereinander hatten.
Werkzeuggebrauch

In Europa sind die Epoche des Moustériens und die in Levalloistechnik hergestellten Steinwerkzeuge mit den Neandertalern assoziiert. Universalwerkzeug zum Schneiden und Schaben war für die „klassischen“ Neandertaler der Würm- bzw. Weichsel-Kaltzeit das Keilmesser, zugleich typologische Leitform des Micoquien (heute: „Keilmesser-Gruppen“). Ein modernes Pendant dieser Gerätform, die sowohl zum Schneiden als auch Schaben eingesetzt wurde, ist bei den Eskimos mit dem Ulu überliefert. Die Fundstellen von Werkzeugen, die während des Moustériens genutzt wurden, befinden sich oft nur fünf bis sechs Kilometer entfernt vom natürlichen Vorkommen des Gesteins, aus denen sie gefertigt wurden; aus diesem Befund wurde auf ein relativ kleines Aufenthaltsgebiet dieser Neandertaler-Gruppen geschlossen. Andere Gruppen zogen über große Entfernungen zu Feuersteinvorkommen, um sich dort mit dem Rohmaterial zu versorgen. Manche Fundplätze waren Eckpunkte eines Streifgebietes von mehr als 100 km Durchmesser.
Wie durch die 1946 gefundene, rund 120.000 Jahre alte Stoßlanze von Lehringen belegt ist, benutzten Neandertaler hölzerne Waffen (Spieße) zum Erlegen von Großwild. Seit 1994 wurden im Tagebau Schöningen zudem acht Speere aus der Holstein-Warmzeit gefunden (Schöninger Speere), die rund 300.000 Jahre alt sind und als Wurfspeere interpretiert werden. Experimente mit Wurfspeeren zeigen, dass diese bis auf eine Entfernung von 20 Meter und mehr treffsicher eingesetzt werden können. Experimente aus 5 Meter Entfernung ergaben durchschnittliche Eindringtiefen von 23,8 cm bei mittleren Auftreffgeschwindigkeiten von 83 km/h und einer Durchschlagskraft von 25,9 N. Die Bewehrung von Lanzen mit Blattspitzen ist für die späten Neandertaler sehr wahrscheinlich, auch die Schäftung von hölzernen Waffen mit Levallois-Spitzen ist in mehreren Fällen bewiesen. In der Fundstätte Poggetti Vecchi in der Provinz Grosseto (Italien) wurden mehrere Dutzend 171.000 Jahre alte, im Feuer gehärtete Grabstöcke geborgen, die zumeist aus dem Holz von Buchsbaum (Buxus sempervirens), aber auch aus Eichen, Wacholder und Eschen hergestellt worden waren. Auch die Benutzung von Zahnstochern gilt als gesichert.
Neandertaler vom Fundplatz Königsaue am Ascherslebener See (Harzvorland) verwendeten Birkenpech zum Einkleben von Steinartefakten in hölzerne Schäfte, ein weiterer Fund von Birkenpech an einem rund 50.000 Jahre alten Steinartefakt wurde aus den Niederlanden berichtet. Für die Destillation des Pechs aus Birkenrinde durch Verschwelung unter Luftabschluss ist eine längere Zeit gleichbleibende Temperatur von etwa 350 °C nötig; jedoch könnte auch ein weniger aufwändiges Verfahren (ohne Luftabschluss) zum Erfolg geführt haben.
Die im Südwesten Frankreichs in den Ausgrabungsstätten Abri Peyrony und Pech-de-l'Azé entdeckten Schleifwerkzeuge (Glätter) aus Hirschknochen, die auf ein Alter von bis zu 50.000 Jahren datiert wurden, ähneln dagegen den bis heute verwendeten Glätthölzern (Lissoirs), mit denen Leder bearbeitet wird. Diese bisher ältesten Spezialwerkzeuge Europas dienten durch Schaben, Schleifen und Polieren dem Weichmachen des Leders und erhöhten die Wasserbeständigkeit. Nach Ansicht der Forscher des Max-Planck-Institut für evolutionäre Anthropologie in Leipzig wäre das möglicherweise ein Beleg dafür, dass die Neandertaler bereits eine eigene Technologie hatten, deren Entstehen bisher dem modernen Menschen zugeschrieben wurde, die dieser jedoch auch vom Neandertaler übernommen haben könnte. Bei ihrer Einwanderung nach Europa kannten sie nur spitze Knochenwerkzeuge, stellten kurze Zeit später aber Lissoirs her.
Jagd
Ergänzend zu den Steinwerkzeugen gibt es Hinweise auf Holzbearbeitung und für die Verwendung von hölzernen Lanzen, so z. B. eine 2,45 Meter lange Eibenholzlanze, mit der vor 120.000 Jahren Waldelefanten erlegt wurden. Man benutzte auch hölzerne, angespitzte Wurfspeere, gelegentlich versehen mit Steinspitzen.
Die Art der verwendeten Steinwerkzeuge und Waffen richtete sich nach Verfügbarkeit des Rohmaterials, nach Überlieferung und individueller Fertigkeit. Es gab Neandertaler, die vorzugsweise in der Nähe von Steinbrüchen siedelten; andere zogen über große Entfernungen zu Feuersteinvorkommen, um sich dort mit dem Rohmaterial zu versorgen. So hatten Gruppen, die sich an den Kratern der Osteifelvulkane aufhielten, Werkzeuge aus Feuerstein dabei, dessen nächstes Vorkommen im Maasgebiet lag (bei Aachen und Maastricht), aber auch so genannten baltischen Feuerstein aus dem Ruhrgebiet. Diese Fundplätze waren Eckpunkte eines Streifgebietes von mehr als 100 km Durchmesser. Manche Fundstellen belegen die Bejagung von Einzeltieren, an anderen Orten gibt es Hinweise auf Massenjagd: Am Fundort Salzgitter-Lebenstedt hatten Neandertaler spezielle Jagdlager aufgeschlagen; hier wurden Jagdbeutereste mit Schlachtspuren von 86 Rentieren gefunden und tausende Steingeräte. Die Jagdperiode lässt sich anhand untersuchter Zähne und der Geweihentwicklung auf den Herbst festlegen. Mittelgroße Säugetiere wie Pferd, Wildesel und Ren wurden oft einzeln erlegt und zerlegt, die Teile zu den Wohnplätzen geschafft. Auf der heutigen Insel Jersey wurden Großsäuger (Elefant, Nashorn) über Kalkklippen getrieben.
Im Bereich der Fundstätte Neumark-Nord 1 (Sachsen-Anhalt) wurden in den 1980er- und 1990er-Jahren beim Abbau von Braunkohle die fossilen Überreste von mindestens 70 Waldelefanten geborgen. Die rund 125.000 Jahre alten Knochen waren in feinkörnigem Seesediment eingebettet, gut erhalten und stammten zum einen fast ausschließlich von erwachsenen Tieren. Zum anderen fiel auf, dass es sehr viele männliche Elefanten waren, was von keinem anderen „Elefantenfriedhof“ bekannt ist. 2023 berichtete ein Forschergruppe um Sabine Gaudzinski-Windheuser, dass man an zahlreichen der mehr als 3000 untersuchten Knochen Schnittspuren entdeckt habe, die gehäuft an Stellen vorkommen, die typisch für das Zerlegen von Jagdbeute sind; gestützt wurde diese Interpretation durch den Fehlen von Bissspuren von Aasfressern. Den Forschenden zufolge wurde der Ort über mindestens 2000 Jahre hinweg aufgesucht. Die Häufigkeit von männlichen Elefanten deute darauf hin, dass diese – zumeist einzelgängerischen Individuen – leichter erbeutet werden konnten als in Gruppen umherziehende Weibchen. Bereits 2018 hatte die gleiche Forschergruppe nachgewiesen, dass die fossilen Knochen zweier Damhirsche aus der Fundstätte Neumark-Nord Löcher aufweisen, die vermutlich aus geringer Entfernung durch Speere verursacht wurden. Anhand eines experimentellen ballistischen Versuchsaufbaus wurde „die Nutzung eines hölzernen Speers in Aufwärtsbewegung, der mit geringer Geschwindigkeit eingesetzt wurde“, rekonstruiert; dies deute darauf hin, „dass sich Neandertaler den Tieren bis auf sehr kurze Distanz näherten und den Speer als Stoß- und nicht als Wurfwaffe verwendeten. Eine solche konfrontative Art der Jagd erforderte sorgfältige Planung, Tarnung sowie ein enges Zusammenspiel zwischen den einzelnen Jägern.“
Kleidung
Die Neandertaler fertigten vermutlich als erste Menschenart Kleidung an, jedoch sind bislang bei ihnen – anders als bei den Cro-Magnon-Menschen – keine Hinweise auf die Herstellung und Verwendung von Nadeln entdeckt worden. Im Abri du Maras (am Ende der Gorges de l’Ardèche, Département Ardèche, Frankreich) wurden allerdings in unmittelbarer Nähe von Steingeräten zu Fäden verdrillte Pflanzenfasern entdeckt, die in solchem Zustand in der Natur nicht vorkommen, 40.000 bis 50.000 Jahre alt sind und aufgrund dieser Datierung dem Neandertaler zugeschrieben wurden. Aus Neumark-Nord, einer ca. 200 000 Jahre alten Fundstelle an einem ehemaligen Seeufer bei Frankleben in Sachsen-Anhalt, stammt ein Steingerät mit anhaftenden Resten von Eichensäure in einer Konzentration, die nicht natürlich auftreten kann und deshalb als ein Hinweis auf das Gerben von Tierhäuten gedeutet wird. In der Fundstelle am Strand von Le Rozel wurden zahlreiche Fußabdrücke gefunden, von denen einige vermutlich von leichtem Schuhwerk stammen.
Auch aus Modellrechnungen wurde abgeleitet, dass die Neandertaler vermutlich Kleidung angefertigt und getragen haben. Den Berechnungen zufolge hätte ein Neandertaler, dessen Körpergewicht 80 kg betrug, während der damaligen Kälteperioden zusätzlich 50 kg Unterhautfettgewebe bilden müssen, um der Kälte unbekleidet zu widerstehen. Ian Tattersall kommentierte dies so: „Gebaut wie ein Sumo-Ringer zu sein, ist kaum das, was man als ideale Anpassung an eine Lebensführung als Jäger ansehen kann.“
Sprache
Im israelischen Karmelgebirge wurde im Jahre 1983 in der Kebara-Höhle das bisher einzige Zungenbein eines Neandertalers entdeckt. Es entspricht dem der modernen Menschen und gilt als wichtigstes Indiz dafür, dass die Neandertaler die anatomische Voraussetzung für die Fähigkeit zum Sprechen besaßen. Durch paläogenetische Untersuchungen wurde im Oktober 2007 ferner festgestellt, dass die Neandertaler über das gleiche FOXP2-Gen wie der moderne Mensch verfügten. Das für die Entwicklung der Sprache als wichtig gedeutete FOXP2-Gen wurde durch DNA-Sequenzierung aus Knochen eines Neandertalers, die in einer spanischen Höhle gefunden wurden, isoliert und analysiert. Eine Rekonstruktion der Schallübertragung zum Innenohr bei fünf Neandertaler-Schädeln ergab zudem, dass sich die Breite des Frequenzbands von Homo sapiens und Neandertalern kaum unterscheidet; zugleich wurden aber erhebliche Unterschiede zwischen Neandertalern und deren Vorläufern (Homo heidelbergensis) nachgewiesen. Daraus wurde geschlossen, dass Neandertaler gesprochene Sprache ähnlich gut hören konnten wie heute lebende Menschen. Auch wenn es noch weiterer Indizien bedarf, gibt es keinen Grund anzunehmen, dass der Neandertaler nicht sprechen konnte.Katarina Harvati und Maria Kirady spekulieren, dass es wohl keine „Gemeinsprache“ gab, die von allen Neandertalern verstanden wurde, und dass wahrscheinlich die Idiome anders strukturiert waren als beim Homo sapiens.
Körperschmuck, symbolisches Denken

Im Südosten Spaniens wurden in zwei Höhlen mehrere Muschelschalen entdeckt, die im Bereich ihres Wirbels (ohne Zutun ihrer Sammler) 5 mm große Löcher aufweisen und laut Radiokohlenstoffdatierung 45.000 bis 50.000 Jahre alt sind; beide Höhlen sind als Aufenthaltsorte von Neandertalern bekannt. Die Schale einer Großen Pilgermuschel aus der Cueva Antón ist auf ihrer Außenseite mit orangefarbenem Pigment bemalt, mehrere Muschelschalen aus der Cueva de los Aviones weisen rote, gelbe und orangefarbene Pigmente auf. In ihrer Nähe wurden zudem weitere Reste von roter und gelber Farbe gefunden. Diese Funde wurden als Beleg dafür gedeutet, dass die Urheber der Fundstücke die Muschelschalen und die Pigmente „in einer ästhetischen und vermutlich symbolischen“ Weise – möglicherweise an einem Halsband befestigt – verwendeten. Einer 2018 publizierten Studie zufolge sind die Funde aus der Cueva de los Aviones laut Uran-Thorium-Datierung sogar 115.000 bis 120.000 Jahre alt.
Ebenfalls in Spanien – nördlich von Madrid – wurden in einer Höhle (Cueva Des-Cubierta) 35 mit Hörnern oder Geweihen bewehrte Schädelteile von großen Pflanzenfressern wie Steppenbisons, Auerochsen, Rothirschen, Rehen und Steppennashörnern gefunden, die dort offenbar als Jagdtrophäen aufbewahrt worden sind.
Auch die in Frankreich gefundene Maske von La Roche-Cotard wird einem Neandertaler zugeschrieben. Ferner wurden in Frankreich, bei Ausgrabungen in Pech de l’Azé, manganhaltige Pigmentklumpen gefunden, die auf eine Körperbemalung der Neandertaler schließen lassen. Die meisten Funde von Farbpigmenten stammen aus der Epoche vor 60.000 bis 40.000 Jahren; der älteste Fund – roter Ocker, dessen Verwendung ungeklärt ist – stammt aus Maastricht-Belvédère und ist mit 250.000 bis 200.000 Jahren ähnlich alt wie Pigmentfunde aus Afrika, die dem frühen Homo sapiens zugeschrieben werden.
In der italienischen Grotta di Fumane (Höhle von Fumane), 18 km nordwestlich von Verona, fanden sich 44.000 Jahre alte Hinweise auf die Entfernung großer Federn von Vogelarten, die nicht verzehrt wurden, wie etwa von Bartgeiern oder Rotfußfalken. Auch entdeckte man Anzeichen für Körperbemalung.
Acht aus den Grabungen von Krapina (Kroatien) erhaltene Klauen von Seeadlern wurden im Jahr 2015 als Teile von Schmuck gewertet: Die Untersuchung mit Lichtmikroskopen schloss bei eingekerbten und polierten Stellen an den Klauen natürliche Herkunft oder zufällige Einwirkungen aus und führte zu dem Schluss, dass die Klauen als dekorative Teile einer Halskette verwendet wurden. Daraus folgerten die Autoren auf eine symbolische Nutzung von Gegenständen durch Neandertaler in Europa vor 130.000 Jahren und somit vor ihrem Kontakt mit modernen Menschen. Unterstützt wurde diese Interpretation 2019 durch den Nachweis von Schnittspuren an Flügelknochen von Steinadlern (an Knochen, die kaum Fleisch ansetzen) aus anderen Fundstellen in Mittel- und Westeuropa, was als Beleg für eine sorgfältige Abtrennung der Federn von den Knochen bewertet wurde. Dies gab den Autoren Anlass für die Vermutung, dass die Federn als Schmuck gedient haben könnten. Schnittspuren fand man auch an einem Zehen-Knochen aus der Foradada-Höhle in Calafell (Spanien), der vermutlich einem Spanischen Kaiseradler zuzuschreiben ist.
Auch „ein winkelartiges Muster aus sechs Kerben“ auf dem Riesenhirsch-Knochen aus der Einhornhöhle im Harz wurde vor mindestens 51.000 Jahren von einem Neandertaler geritzt.
Höhlenmalerei
Die ältesten aus Europa bekannten Höhlenmalereien sind rund 65.000 Jahre alt. Ein internationales Forscherteam unter der Leitung von Dirk Hoffmann vom Max-Planck-Institut für evolutionäre Anthropologie berichtete im Jahr 2018, dass Neandertaler in Europa schon mehr als 20.000 Jahre vor der Ankunft anatomisch moderner Menschen in Europa vor 40.000 Jahren Höhlenkunst geschaffen haben. Die Forscher hatten mit Hilfe der Uran-Thorium-Datierung 60 Proben von Karbonat-Krusten auf den Farbpigmenten von Malereien in drei Höhlen in Spanien analysiert: aus der Cueva de La Pasiega in der Gemeinde Puente Viesgo, der Maltravieso-Höhle in der Gemeinde Cáceres und der Höhle von Ardales (in der Gemeinde Ardales, Südspanien). „Sie enthalten meist rote, manchmal auch schwarze Malereien, die Tiergruppen, Punkte, geometrische Zeichen sowie positive und negative Handabdrücke und auch Felsritzungen umfassen.“ Eine weitere wissenschaftliche Veröffentlichung führte später allerdings an, dass die Datierung falsch sein könnte und dass bei manchen für Malereien gehaltene Farbschichten eine natürlich-geologische Ursache nicht auszuschließen sei. Im Jahr 2021 veröffentlichte Ergebnisse einer Studie widerlegten diese Theorie. Mittels Röntgenspektroskopie, Mikro-Raman-Spektroskopie und Röntgenbeugung konnte eine natürlich-geologische Ursache ausgeschlossen und die Farbschichten als Höhlenmalerei identifiziert werden.
Lange ging die Wissenschaft davon aus, dass in den Malereien Männer ihre Jagderfahrungen künstlerisch umgesetzt hatten, doch gab es dafür keine Beweise. Der Archäologe Dean Snow von der Pennsylvania State University analysierte Handabdrücke aus acht französischen und spanischen Steinzeithöhlen, darunter der El-Castillo-Höhle, und fand heraus: Etwa drei Viertel aller farbigen Hände stammen von Frauen, und es finden sich auch zahlreiche Handabdrücke von Kindern und Jugendlichen.
Räumlich differenzierte Nutzung von Höhlen
In dem als logistisches Basislager angesprochenen Felsüberhang Bombrini im Nordwesten Italiens lebten Neandertaler offenbar auf drei Ebenen. Jede dieser Ebenen enthielt Artefakte, die es ermöglichten, auf eine Aufteilung in Räume zu schließen, die zur Schlachtung, zum Wohnen und zur Werkzeugherstellung dienten.
In einer Höhle bei Bruniquel in Südfrankreich wurden zahlreiche Fragmente von gezielt abgebrochenen, mehr als 400 Stalagmiten entdeckt, die zu zwei Ringen von 6,70 bzw. 4,50 Meter Umfang angeordnet sind (Steinkreise in der Höhle von Bruniquel). Diese Konstruktionen wurden vor 176.500 ± 2100 Jahren errichtet, rund 330 Meter entfernt vom Höhleneingang. Die Funktion dieser Ringe ist ungeklärt, jedoch wurde ein Daumen-großes, verbranntes Stück Knochen entdeckt, woraus geschlossen wurde, dass hier möglicherweise Nahrung zubereitet wurde. Dies kann jedoch nicht erklären, warum die Neandertaler einen derart abgelegenen, schwer zu erreichenden Ort mit so großem Aufwand ausstatteten.
Bestattungen


Aufgrund zumindest vereinzelter Bestattungen ihrer Toten sowohl in Europa als auch im Nahen Osten und dem Ablegen von Toten in Höhlen ist Homo neanderthalensis neben Homo sapiens die fossil am besten überlieferte Art der Hominini. „Der Verstorbene wurde zumeist in Rückenlage oder auch in Hockerstellung – also auf der Seite liegend mit angezogenen Beinen – in das Grab gebettet. Farbpigmentreste von Rötel und Ocker identifizierte man in den Gräbern bei La Ferrassie, Spy und La Chapelle-aux-Saints. Welche Bedeutung Farben bei Beerdigungen von Neandertalern zukam und auf welche kultischen Praktiken sich der Gebrauch von Naturpigmenten zurückführen lässt, ist unbekannt.“ Ein großer Unterschied zwischen den Gräbern der Neandertaler und der Cro-Magnon-Menschen besteht vor allem in den Grabbeigaben:
„Jungpaläolithische Gräber waren häufig sehr komplex mit reichgeschmückten Toten und zahlreichen Grabbeigaben. Entsprechend gedeutete Gegenstände in Moustérien-Gräbern waren dagegen meist alltägliche Gegenstände wie Steinwerkzeuge und einzelne Tierknochen. Diese könnten als Ausrüstung und zur Versorgung im späteren Leben gemeint gewesen sein, es wäre aber auch denkbar, daß sie als allgegenwärtige Gegenstände des Wohnraumes eher zufällig mit in das Grab gelangten. Es gibt nur wenige Dinge in Moustérien-Gräbern, deren Deutung als ‚Grabbeigabe‘ einer kritischen Analyse standhält.“
Bereits 1945 wurde aber beispielsweise das Grab eines ungefähr neun Jahre alten Neandertaler-Jungen beschrieben, das in der Teschik-Tasch-Höhle in Usbekistan entdeckt worden war; das Skelett des Kindes lag dort rund 70.000 Jahre lang umsäumt von Steinbock-Hörnern.
Ungefähr gleich alt sind mehrere Neandertaler-Grabstätten in der Shanidar-Höhle (Irak). In Grab IV wurde eine ungewöhnlich hohe Konzentration von Blütenpollen nachgewiesen, was gelegentlich als Beleg „für Schamanismus und ritualisierte Bestattungen“ interpretiert wurde; die Blüten könnten aber auch von den dort häufig vorkommenden Persischen Rennmäusen in die Höhle verschleppt und in den Bestattungshorizont eingegraben worden sein. Kontrovers als Bestattungen oder Niederlegungen in Gruben diskutierte Neandertaler-Funde gibt es außerdem im Abri La Ferrassie (Südwestfrankreich). Im Jahre 2011 wurden Bestattungsbefunde aus der spanischen Höhle Sima de las Palomas del Cabezo Gordo bekannt. Auch der sogenannte Alte Mann von La Chapelle wurde aus einer Grube geborgen, „deren Füllung sich farblich eindeutig vom umgebenden Sediment unterscheidet.“
Knochen von Höhlenbären in der Schweizer Drachenlochhöhle, die zwischen Steinplatten angeordnet waren, waren die Ursache für einen den Neandertalern unterstellten Bärenkult. Die Felsen können freilich auch ohne menschliche Einwirkung von der Höhlendecke herabgefallen, die „ausgerichtet“ wirkende Anordnung der Funde durch Wassereinwirkung erfolgt sein. Zumal es keine weiteren Belege für einen so frühen Bärenkult gibt (etwa Ritualgegenstände, vergemeinschaftete Bestattungen etc.) und existierende Bärenkulte sehr komplex sind, wird dessen Existenz heute als wenig wahrscheinlich bzw. widerlegt bewertet.
Fortpflanzung und Bevölkerungsdichte
Genetische Analysen von Zahnfunden in der spanischen El-Sidron-Höhle deuten auf ein patrilokales Fortpflanzungsverhalten der Neandertaler hin. Carles Lalueza-Fox vom Institut für Evolutionsbiologie in Barcelona, der diese Analysen anhand mitochondrialer DNA (mtDNA) an einer 12-köpfigen familiär verwandten Neandertalergruppe durchgeführt hat, deutet dies als eine soziale Praxis der Neandertaler, wie sie auch bei modernen Jäger- und Sammler-Kulturen vorkommt, nämlich, dass die Frauen ihre ursprünglichen Gruppen verließen, während die Männer in der Gruppe des Vaters verblieben. Ob damit auf eine durchgängig patrilineare Sozialpraxis der Neandertaler geschlossen werden kann, ist allerdings noch nicht abschließend geklärt. Die auf Basis der mtDNA analysierten Abstammungslinien innerhalb der Gruppe lassen ferner eine Geburtshäufigkeit der Neandertaler von ca. drei Jahren plausibel erscheinen.
Anhand der ausschließlich von der Mutter auf die Kinder übertragenen mtDNA von fünf Neandertalern wurde 2009 berechnet, dass vor rund 70.000 bis 40.000 Jahren allenfalls 3500 weibliche Neandertaler gleichzeitig gelebt haben. Wie aussagekräftig dieser Schätzwert ist, blieb allerdings umstritten. Zum einen wurde aus ihm abgeleitet, dass die Gesamtpopulation zu einem bestimmten Zeitpunkt dieser Spätphase der Neandertaler nur bei 7000 Individuen gelegen habe; zugleich wurde in einem Begleitartikel zur Studie in der Zeitschrift Science aber auf Modellrechnungen zur heutigen Einwohnerschaft in Schweden verwiesen, wo rund neun Millionen Menschen leben. Ein vergleichbares Vorgehen wie beim Neandertaler würde für die heutige schwedische Homo-sapiens-Population aber nur auf 100.000 Individuen kommen; daher könne die tatsächliche Zahl weiblicher Neandertaler in der genannten Epoche durchaus 70.000 betragen haben.
Jean-Jacques Hublin vom Max-Planck-Institut für evolutionäre Anthropologie kommt hingegen 2018 zu dem Schluss, dass es „selbst in Zeiten ihrer größten Verbreitung“ nicht mehr „als geschätzte 10.000 ‚Neandertal-Europäer‘“ zugleich gegeben habe, wobei die Größe der einzelnen Gruppen „höchstens 50 bis 60 Frauen und Männer“ umfasst habe.
Beispiele für bedeutende Fundstätten
Fundplätze auf der Halbinsel Krim

Fundplätze auf der Halbinsel Krim weisen zum Teil Kulturschichten von der Eem-Warmzeit (125.000 Jahre vor heute) bis zum Aussterben der Neandertaler vor etwa 30.000 Jahren auf. Demnach blieb die Werkzeugkultur über mehrere zehntausend Jahre weitgehend unverändert: flache Klingen, die meist beidseitig durch Oberflächenretuschen über lange Zeit funktionstüchtig gehalten wurden. Sie befanden sich in seitlichen Griffschäften aus Holz- oder Knochen und wurden zum Teil im montierten Zustand nachretuschiert. Diese „Ak-Kaya-Industrie“ genannte Kultur ähnelt der ebenfalls dem Neandertaler zugeschriebenen Industrie des Micoquien Mitteleuropas. Mit dem Absinken der Temperaturen zum ersten Maximum der letzten Eiszeit vor etwa 60.000 Jahren änderte sich die Kultur: Es wurden nun durch Abschlagstechnik Werkzeuge aus Feuerstein erzeugt, die nach Abnutzung weggeworfen und nicht nachretuschiert wurden. Die Kultur der späten Neandertaler ähnelte daher dem Aurignacien des Homo sapiens in Mitteleuropa, obwohl dieser auf der Krim erst vor 30.000 Jahren nachgewiesen ist. Die Neandertaler hatten demnach auf der Krim wichtige Innovationen des modernen Menschen gleichsam vorweggenommen.
Die zahlreichen Knochenfunde von Wildeseln am Fundplatz Kabazi machen deutlich, dass die Neandertaler fähig waren, bei Beutezügen planmäßig vorzugehen. Die Funde wurden dahingehend interpretiert, dass ganze Familien bzw. Herden von Eseln mit Eltern- und Jungtieren erbeutet wurden, während diese am nahen Flusslauf tranken. Die Beute wurde an Ort und Stelle zerlegt, aber wesentliche Teile der Tiere wurden im Stück abtransportiert und an anderer Stelle zerteilt, zubereitet und verzehrt. Nachgewiesen wurde auch eine über unterschiedliche Lagerplätze verteilte Arbeitsteilung: So gab es Lagerplätze, wo das Wild zerlegt und die Steinwerkzeuge hergestellt wurden, sowie andere, wo offensichtlich länger gewohnt und häufiger verzehrt wurde. Eine deutliche planmäßige Arbeitsteilung und Organisation, eine jahreszeitlich ausgerichtete Spezialisierung auf einzelne Tierarten und saisonale Lagerplätze bezogen auf die ganze Gruppe konnten ausgemacht werden. Die Forscher gewannen den Eindruck von mittelpaläolithischen Menschen, die gewisse jungpaläolithische Errungenschaften bereits entwickelt hatten, andere aber noch nicht kannten: Nicht nachgewiesen wurden beispielsweise typische jungpaläolithische Merkmale wie besondere Geweih- und Knochenbearbeitungen sowie Werkzeuge wie Stichel und Kratzer.
Am Fundort Kiik-Koba, einer großen Höhle im Krimgebirge (44°57' N, 34°21' E), wurde ein Feuerstein mit Ritzungen geborgen, dessen Fundschicht mittels kalibrierter C14-Datierung ein Alter von 35.000 bis 37.000 Jahren (35 bis 37 cal kyr BP) datiert wurde; die Ritzungen wurden als möglicherweise absichtsvoll gestaltete, figürliche Gravur interpretiert.
Fundplätze in Deutschland und Österreich

Der bekannteste Fundplatz in Deutschland ist das Neandertal, wo aber nur wenige Steinwerkzeuge gefunden wurden, die zudem nicht in unmittelbarem Fundzusammenhang mit dem namensgebenden Fossil Neandertal 1 standen.
Ein bedeutender Fundort in Deutschland ist hingegen die Balver Höhle in Westfalen, weil sie in der ersten Hälfte der Weichsel-Kaltzeit vor 100.000 bis 40.000 Jahren immer wieder von Neandertalern aufgesucht wurde. Im Fundmaterial aus der Balver Höhle konnten neben zahlreichen Steinartefakten viele Geräte aus Knochen und Mammutelfenbein identifiziert werden. Das Sediment der Höhle war zudem durchsetzt mit den Knochen von Mammuts, darunter vor allem von Kälbern und Jungtieren; es wird vermutet, dass die sehr große Anzahl von Tieren im Umfeld der Höhle erlegt worden ist. In der Gudenushöhle (Kleines Kremstal, Niederösterreich) deutet die untere, 70.000 Jahre alte Kulturschicht auf Jagd von Mammuts, Wollnashörnern, Rentieren, Wildpferden und Höhlenbären hin. Aufgrund der häufigen Kopf- und Armverletzungen an Neandertaler-Skeletten schließt man darauf, dass das Großwild mit Nahwaffen gejagt wurde, wie auch der Fund einer Holzlanzenspitze belegt.
Mögliche Seefahrt (in Griechenland)
Hinweise auf frühe seefahrende Neandertaler wurden im östlichen Mittelmeerraum gefunden, wo die Neandertaler und deren Vorfahren (Homo heidelbergensis) seit rund 300.000 Jahren ansässig waren. Ihre typischen Moustérien-Steinwerkzeuge wurden jedoch nicht nur auf dem griechischen Festland gefunden, sondern – datiert auf ein Alter von mindestens 110.000 Jahren – auch auf den griechischen Inseln Lefkada, Kefalonia und Zakynthos. Mit Ausnahme von Lefkada – während der Eiszeiten bei einem um bis zu 120 Meter tieferen Meeresspiegel eine Halbinsel des griechischen Festlands – bildeten Kefalonia und Zakynthos einschließlich Ithaka in diesen Zeiten eine einzige große Insel. Sie war von mindestens 180 Meter tiefem Wasser umgeben und konnte vermutlich nur mit Wasserfahrzeugen erreicht werden; der Abstand zum Festland betrug seinerzeit etwa 5 bis 7,5 Kilometer zur Südspitze der Halbinsel von Lefkada.
Bereits in den Jahren 2008 und 2009 hatten Forscher um Thomas Strasser vom Providence College in der Schlucht des Megalopotamos auf Kreta, oberhalb des Palmenstrandes von Preveli, 130.000 Jahre alte Steinwerkzeuge gefunden; auch diese Werkzeuge stammen aus einer Epoche, in der Homo sapiens in Europa noch nicht ansässig war. Kreta ist seit etwa 5,3 Millionen Jahren vollständig von Wasser umgeben, das nächste Land war auch während der Eiszeiten rund 40 Kilometer entfernt. Strasser ordnet die Funde auf Kreta allerdings nicht dem Homo neanderthalensis, sondern dem Homo heidelbergensis oder dem Homo erectus zu.
Auch auf Naxos gibt es Fundplätze von Steingerät aus dem Mittelpaläolithikum; ob diese Insel während der Eiszeiten zumindest zeitweise vom Festland aus trockenen Fußes erreichbar war, ist ungeklärt. 2019 wurde für die Funde ein Alter von rund 200.000 Jahren ausgewiesen.
In Asien konnte Homo erectus die Insel Flores nur besiedeln, nachdem er vor rund einer Million Jahren mit Wasserfahrzeugen mehrere, auch während der Eiszeiten bestehende Wasserstraßen zwischen den Nachbarinseln überwunden hatte (vergl. Homo floresiensis).
Aussterben
Die Gründe für das Aussterben der Neandertaler sind umstritten. Archäologische Belege für kriegerische Handlungen zwischen Neandertalern und anatomisch modernen Menschen gibt es ebenso wenig wie Anzeichen für eine sehr rasche Verdrängung der Neandertaler durch den anatomisch modernen Menschen. Im Gegenteil: Bard et al. (2020) wiesen nach, dass Neandertaler und anatomisch moderner Mensch für etwa 3960 ± 710 Jahre gemeinsam in Europa existierten.
Zahlreiche Hypothesen wurden aufgestellt, um das Verschwinden der Neandertaler zu erklären. In einer 2021 veröffentlichten Übersichtsarbeit wurden diese Hypothesen nach folgenden Ursachen gruppiert:
- Wettbewerb um begrenzte Ressourcen. Je nach Auffassung der einzelnen Forscher werden folgende Eigenschaften des anatomisch modernen Menschen für seine Überlegenheit in den Vordergrund gestellt, und zwar
- morphologische Merkmale
- kognitive Merkmale
- technologische Merkmale
- soziale Merkmale
- ökonomische Merkmale.
- Interne demografische Dynamik der Neandertaler-Populationen:
- Selbst ohne Konkurrenz durch den anatomisch modernen Menschen sei die Neandertaler-Population zu klein gewesen, um langfristig bestehen zu können.
- Die geringe Größe der lokalen Teilpopulationen und deren zu geringe Vernetzung mit anderen Teilpopulationen habe die Neandertaler anfällig für Inzucht gemacht und zu Inzuchtdepression infolge des fehlenden Allee-Effekts geführt.
- Veränderungen in der Umwelt:
- das Klima wurde instabiler (u. a. im Rahmen des als H4 bezeichneten Heinrich-Ereignisses vor ca. 40000 Jahren).
- es kam zu klimatischen Extremsituationen nach dem Ausbruch eines Vulkans in den Phlegräischen Feldern der süditalienischen Region Kampanien (vergl. Kampanischer Ignimbrit).
- die anatomisch modernen Menschen brachten Krankheitserreger mit sich, gegen die sie immun waren, nicht aber die Neandertaler.
Einige dieser Hypothesen wurden in jüngerer Zeit durch Forschungsarbeiten infrage gestellt. Beispielsweise veröffentlichte ein Wissenschaftlerteam im Jahr 2012 eine Studie, der zufolge das vor 40.000 bis 30.000 Jahren allmählich kälter werdende Klima in Europa anscheinend keinen maßgeblichen Einfluss auf das Aussterben der Neandertaler hatte, denn die Hauptvereisung Europas begann erst vor rund 25.000 Jahren und erreichte ihr Maximum vor 20.000 Jahren, als die Neandertaler bereits seit langem ausgestorben waren. Laut einer Studie aus dem Jahr 2018 gab es jedoch vor 44.000 bis 40.000 Jahren zahlreiche Wechsel von ausgeprägten Kaltphasen und minder kalten Interstadialen. Dies habe wiederholt zu regionaler Neandertaler-Entvölkerung und nachfolgend – möglicherweise – zu einer Besiedelung der entvölkerten Regionen durch Homo sapiens geführt.
Erwogen wird ferner, dass die gemeinsame Jagd mit frühen „Wolf-Hunden“ Vorteile für die Cro-Magnon-Menschen brachte. Die US-Archäologin Paola Villa und ihr niederländischer Kollege Wil Roebroeks fanden allerdings im Jahr 2014 in der gesamten Fachliteratur keinerlei archäologische Befunde, die eine kulturelle Überlegenheit des Homo sapiens gegenüber dem Neandertaler belegen; stattdessen vermuten sie eine allmähliche zahlenmäßige Überlegenheit des Homo sapiens. Modellrechnungen belegen nämlich, dass Gruppengröße und Populationsdichte Einfluss auf die kulturelle Komplexität haben können. Der deutsche Paläoanthropologe Friedemann Schrenk vermutet ebenfalls: „Am wahrscheinlichsten erscheint die Theorie des Neandertalers als Fortpflanzungsmuffel. So genannte ‚bottle-neck‘-Situationen, also Bevölkerungsengpässe, waren keine Seltenheit in der Geschichte der Menschheit und könnten daher auch den Neandertaler betroffen haben.“ Eine 2021 bekannt gewordene Umfrage unter mehr als 200 Paläoanthropologen ergab, dass heute die überwiegende Mehrzahl der Forscher populationsbiologische Nachteile der Neandertaler gegenüber dem anatomisch modernen Menschen als Hauptursache für dessen Aussterben ansieht. Der Vergleich von 17.367 protein-codierenden Genen von Neandertalern aus Spanien, Kroatien und Südsibirien – das heißt aus weit voneinander entfernten Regionen Eurasiens – ergab tatsächlich Hinweise auf eine „bemerkenswert niedrige“ (remarkably low) genetische Vielfalt. DNA- und mtDNA-Proben von 15 Fossilien aus der Tschagyrskaja-Höhle, die zwischen 59.000 und 51.000 Jahre alt sind, wiesen laut einer 2022 publizierten Analyse ebenfalls eine extrem niedrige genetische Vielfalt auf. Entscheidend für das Aussterben könnte ferner gewesen sein, dass der anatomisch moderne Mensch früher geschlechtsreif wurde und mehr Nachkommen hatte.
Ferner gibt es archäologische Hinweise darauf, dass sich beispielsweise in der Region Aquitanien – einem Gebiet mit der größten Dichte an Funden beider Populationen – zwischen 55.000 und 35.000 Jahren vor heute die Zahl der Individuen von Homo sapiens verzehnfachte. Vermutlich konnte Homo sapiens aufgrund seiner kulturell tradierten Verhaltensweisen in dichter besiedelten Gebieten besser überleben als die Neandertaler. Statistische Bevölkerungsmodelle zeigen, dass schon Unterschiede von wenigen Prozent bei der Fortpflanzungsrate ausreichen, um in wenigen tausend Jahren zum Aussterben der weniger begünstigten Population zu führen. In einer Übersichtsarbeit nannte Katerina Harvati im Jahr 2010 neben einer höheren Geburtenrate, kürzeren Abständen zwischen zwei Geburten und dadurch entstehenden größeren Gruppen noch weitere Szenarien, die von einzelnen Forschern – in unterschiedlichen Kombinationen – für möglich gehalten werden: So könnten die anatomisch modernen Menschen beispielsweise eine geringere Sterblichkeit, ein größeres Nahrungsspektrum sowie bessere Kleidung oder bessere Unterkünfte während der Kaltzeiten gehabt haben. Auch unterschiedliche Gewohnheiten beim Warenaustausch wurden erwogen.
Verwandtschaft zum modernen Menschen
Historisches
Nachdem sich Ende des 19. Jahrhunderts die Auffassung durchgesetzt hatte, dass der Neandertaler ein Vorläufer des anatomisch modernen Menschen war, begann in Fachkreisen eine bis heute anhaltende Debatte über deren verwandtschaftliche Nähe. Unterschiedliche Meinungen gab es zunächst insbesondere zur Frage, ob die Neandertaler bloß zeitlich und räumlich Vorläufer des Homo sapiens waren oder ob sich der anatomisch moderne Mensch aus ihnen heraus entwickelt habe. Der deutsche Anatom Gustav Schwalbe untersuchte beispielsweise die bis 1906 bekannten Neandertalerfunde (er bezeichnete sie als Homo primigenius, „ursprünglicher Mensch“) und deutete einige Funde als „Zwischenformen zwischen Homo primigenius und sapiens.“ Die vorherrschende Meinung in den 1910er- und 1920er-Jahren wurde hingegen vor allem durch Arthur Keith geprägt sowie durch Marcellin Boule, der die erste wissenschaftliche Beschreibung eines fast vollständigen Neandertaler-Skeletts verfasst hatte; beide gehörten zu den einflussreichsten Paläoanthropologen ihrer Epoche. Ihrer Auffassung nach war der Körperbau des Neandertalers derart „primitiv“, dass er kein direkter Vorfahre des Homo sapiens sein könne. Diese Auffassung war unter anderem einer fehlerhaften Rekonstruktion des Neandertaler-Fundes La Chapelle-aux-Saints 1 durch Marcellin Boule geschuldet, der das Fossil in krummer Haltung, mit verkrümmter Wirbelsäule und eingeknickten Beinen rekonstruiert hatte.
Diese Interpretation änderte sich in den 1930er-Jahren, als Ernst Mayr, George Gaylord Simpson und Theodosius Dobzhansky den Neandertaler als Homo sapiens neanderthalensis mit dem nunmehr Homo sapiens sapiens genannten anatomisch modernen Menschen der gleichen Art zuordneten. Die scheinbar lückenlose Abfolge von Fundstellen beider – nunmehr – Unterarten in Europa wurde dahingehend interpretiert, dass es einen langsamen, graduellen evolutiven Übergang vom Neandertaler zum anatomisch modernen Menschen gegeben habe. So verteidigte beispielsweise auch Aleš Hrdlička 1927 die Hypothese von der „Neandertaler-Phase des Menschen“.Franz Weidenreich bezeichnete 1943 zum wiederholten Male die Neandertaler als „Zwischenstufe“ („intermediate form“) zwischen dem chinesischen Sinanthropus und Homo sapiens, optierte also ebenfalls für einen kontinuierlichen Übergang, und noch 1964 wurde diese Sichtweise von einer großen und prominenten Autorengruppe verteidigt. Zusammen mit ähnlich gelagerten Interpretationen von Funden in Asien ging aus diesen Überlegungen auch die Hypothese vom multiregionalen Ursprung des modernen Menschen hervor.
In den folgenden vier Jahrzehnten sorgten in Europa zahlreiche neu entdeckte Fossilien und die nachfolgende Neuinterpretation früherer Funde dafür, dass die frühen, sogenannten Präsapiens-Fossilien allesamt in die Vorfahrenlinie der Neandertaler gestellt wurden; hierzu gehörten u. a. der Swanscombe-Schädel aus England, der Schädel aus Steinheim in Baden-Württemberg sowie die Schädel aus Tautavel und aus Biache-Saint-Vaast in Frankreich. Bereits 1973 bescheinigte William W. Howells den Neandertalern in seiner umfangreichen Studie Cranial Variation in Man, dass die anatomischen Merkmale ihrer Schädel außerhalb der Variationsbreite des Homo sapiens liegen und sie folglich einer eigenen Art zugeschrieben werden können. Seine Daten ergaben ferner keine Hinweise darauf, dass es in Europa einen gleitenden Übergang von Neandertalern zum anatomisch modernen Menschen gegeben habe. Bis Anfang der 1980er-Jahre setzte sich daher allmählich die Lehrmeinung in der Paläoanthropologie durch, dass es in Europa nur die zum Neandertaler führende Entwicklungslinie gegeben hat und dass es – bezogen auf Homo sapiens – eine separate Entwicklungslinie gibt, die von afrikanischen Fossilien des frühen Jungpleistozäns zu den Cro-Magnon-Menschen im westlichen Eurasien und zu den Menschen der Gegenwart führt. Diese von Günter Bräuer auf dem 1. Internationalen Kongress für Paläoanthropologie in Nizza erstmals formulierte „Afro-europäische Sapiens-Hypothese“ ist heute bekannt als Out-of-Africa-Theorie. Eine kleine Anzahl von Forschern vertrat aber weiterhin das multiregionale Modell, Milford H. Wolpoff und Alan G. Thorne beispielsweise – gestützt u. a. auf morphologische Analysen von Weidenreich – noch im Jahr 2003, während Forscher wie Günter Bräuer zwar davon ausgingen, dass Homo sapiens in Europa die Neandertaler und in Asien andere archaische Arten wie Homo erectus ersetzte („replacement“ statt „continuity“), was ihrer Auffassung nach aber die Möglichkeit von Genfluss zwischen den Arten nicht ausschloss. Die Abkehr vom vermuteten graduellen evolutiven Übergang des Neandertalers zum anatomisch modernen Menschen hatte zur Folge, dass beiden erneut der Status von Arten (Homo neanderthalenis und Homo sapiens) statt Unterarten zuerkannt wurde. Chris Stringer begründete dies 2001 und erneut 2014 beispielsweise damit, dass die Neandertaler zwar eng verwandt mit Homo sapiens seien, jedoch hinreichend viele nur bei ihnen vorkommende anatomische Merkmale aufweisen,Ian Tattersall bekräftigte diese Argumentation aus dem Blickwinkel des morphologischen Artkonzepts im Jahr 2015, während Fiorenzo Facchini (2006) und einige andere Forscher weiterhin eine Klassifizierung als Unterarten bevorzugen.
Svante Pääbo zufolge ist offen, ob angesichts der genetischen Daten die Einordnung von Neandertalern und anatomisch modernen Menschen in zwei biologischen Arten Bestand haben wird, da es keine Artdefinition gebe, „die für alle Gruppen von Tieren oder Hominiden zutrifft.“
Chris Stringer hatte bereits 2012 zusammenfassend festgestellt: „Obwohl die normale Artendefinition durch die unvollständige reproduktive Abgrenzung nicht gegeben ist, ist es zu früh und angesichts der morphologischen großen Unterschiede aus praktischen Gründen noch nicht erforderlich, H. heidelbergensis, H. neanderthalensis und den Denisova-Menschen mit H. sapiens in einer Art zusammenzufassen.“ Zu den besonders großen morphologischen Unterschieden gehören beispielsweise die Gestalt der Gesichter und der Gehirne beider Arten.
Heutige Sichtweisen

Die Verwandtschaft von Neandertalern und anatomisch modernen Menschen (Homo sapiens) gilt als weitestgehend geklärt. Unter Paläoanthropologen herrscht Einvernehmen darüber, dass beide im afrikanischen Homo erectus einen gemeinsamen Vorfahren hatten. Aufgrund von Fossilien- und Werkzeugfunden gilt es als erwiesen, dass Homo erectus Afrika „während der ersten Ausbreitungswelle vor rund 2 Millionen Jahren“ Richtung Levante, Schwarzmeerraum und Georgien sowie möglicherweise über Nordwestafrika Richtung Südspanien verließ. Diese frühe Besiedelung Georgiens ist durch die 1,8 Millionen Jahre alten homininen Fossilien von Dmanissi belegt. Die ältesten, stark fragmentierten Funde in Europa stammen aus Spanien. Diese 1,2 Millionen Jahre alten Funde werden von ihren Entdeckern als Homo antecessor bezeichnet und als Vorfahren der Neandertaler ausgewiesen; diese Deutung ist allerdings stark umstritten und scheint durch jüngste genetische Befunde zur Verwandtschaft des Neandertalers mit dem anatomisch modernen Menschen widerlegt zu sein.
Vor rund 600.000 Jahren kam es nämlich nach Meinung vieler Paläoanthropologen zu einer zweiten Ausbreitungswelle des afrikanischen Homo erectus. In Spanien gefundene Schädel aus jenem Zeitraum lassen beispielsweise für das Gehirn ein Volumen zwischen 1100 cm³ und 1450 cm³ annehmen; das Gehirnvolumen der Fossilienfunde aus der ersten Ausbreitungswelle wird hingegen auf nur wenig über 1000 cm³ geschätzt. Nach dieser zweiten Besiedelung Europas durch Homo erectus entwickelte sich dieser in Europa über die Homo heidelbergensis genannte Zwischenstufe zum Neandertaler, während in Afrika vor ca. 300.000 Jahren – durch Fossilienfunde belegt – aus Homo erectus der sogenannte frühe anatomisch moderne Mensch und aus diesem der heutige Mensch hervorging.

Stärker umstritten ist jedoch, wann sich die zu den Neandertalern führende Entwicklungslinie von der zum heutigen Menschen führenden Entwicklungslinie getrennt hat. Anhand der molekularen Uhr wurde 2010 zunächst eine Zeitspanne zwischen 440.000 und 270.000 Jahren vor heute berechnet. Die „Ganggenauigkeit“ der molekularen Uhr, die solchen Abschätzungen zugrunde liegt, ist allerdings umstritten; häufig weichen die mit Hilfe geologischer – speziell stratigrafischer – Methoden ermittelten Datierungen erheblich von jenen ab, die mit Hilfe der molekularen Uhr ermittelt wurden. Im Falle der Trennung von Neandertaler und Homo sapiens sprach insbesondere die Datierung der erwähnten spanischen Funde, die der zweiten Ausbreitungswelle von Homo erectus zugeschrieben werden, gegen die Berechnungen mit Hilfe der molekularen Uhr. Eine Neuberechnung der Mutationsraten ergab im Jahre 2012 denn auch Hinweise auf eine deutlich frühere Trennung; sie wurde – recht ungenau – in die Zeitspanne zwischen 800.000 und 400.000 Jahren vor heute datiert. Gestützt wird die Datierung unter anderem durch den rund 400.000 Jahre alten Swanscombe-Schädel, dem – obwohl meist noch zu Homo heidelbergensis gestellt – bereits deutliche Merkmale der frühen Neandertaler zugeschrieben wurden.
Die Datierungsversuche mit Hilfe der „molekularen Uhr“ weichen zudem erheblich von Befunden ab, die aus der Analyse von 1200 homininen Backenzähnen abgeleitet wurden, darunter Zähne von allen Arten der Gattung Homo sowie von Paranthropus. Eine spanisch-amerikanische Forschergruppe hatte anhand von Neandertaler-Zähnen und von Zähnen des anatomisch modernen Menschen die Beschaffenheit der Zähne des letzten gemeinsamen Vorfahren beider Populationen rekonstruiert und diese Rekonstruktion mit den Zähnen früher homininer Arten verglichen. In den Proceedings of the National Academy of Sciences berichteten sie im Jahr 2013, dass – abgeleitet unter anderem vom Wandel der Zahngestalt in der Übergangszeitspanne von Homo heidelbergensis zum Neandertaler – sich die Entwicklungslinien von Neandertalern und anatomisch modernen Menschen ihren Berechnungen zufolge bereits vor 1 Million Jahre trennten. 2019 wurde diese Argumentation in einer weiteren Studie bekräftigt und 800.000 Jahre als Mindestalter für den letzten gemeinsamen Vorfahren berechnet.
Sämtliche Datierungen ergeben jedoch, dass die mitteleuropäischen Populationen von Homo erectus / Homo heidelbergensis und des Neandertalers bis zur Einwanderung des – aus Afrika kommenden – anatomisch modernen Menschen vor rund 40.000 bis 45.000 Jahren mehrere hunderttausend Jahre räumlich getrennt von Homo sapiens lebten. Zwei dem Homo sapiens zugeschriebene Milchzähne aus der Grotta del Cavallo in Apulien wurden auf ein Alter von 45.000 bis 43.000 Jahren vor heute (cal BP) datiert; sie sind der älteste Beleg für den Aufenthalt des Homo sapiens in Europa und belegen zugleich, dass Homo sapiens und Homo neanderthalensis einige tausend Jahre denselben Kontinent besiedelten.
Vermischung anatomischer Merkmale
Das enge Nebeneinander von Fundstellen der Neandertaler und der anatomisch modernen Menschen in der Levante belegt, „dass die beiden Hominidengruppen mindestens 60.000 Jahre ohne Probleme nebeneinander existiert haben.“ Das zu Homo sapiens gehörende Schädeldach Manot 1 aus der Manot-Höhle in Israel wurde beispielsweise auf ein Alter von 54.700 ± 5.500 Jahren (cal BP) datiert, in eine Epoche also, aus der in dieser Region auch diverse Neandertaler-Funde bekannt sind. Daher war die Frage naheliegend, ob trotz der langen Separierung beider Gruppen noch gemeinsame fruchtbare Nachkommen in Europa möglich gewesen sein könnten. Gestützt wurde diese Vermutung zudem durch einen Erfahrungswert, dem zufolge eine vollständige reproduktive Isolation bei mittelgroßen Säugetieren im Allgemeinen mindestens 1,4 Millionen Jahre dauert.
Tatsächlich wurden einige europäische Neandertaler-Funde aufgrund anatomischer Merkmale als Mischlinge interpretiert. Vertreter der „Vermischungshypothese“ war vor allem der US-amerikanische Forscher Erik Trinkaus, der, im Widerspruch zur vorherrschenden Lehrmeinung, nicht nur für Afrika, sondern auch für Asien einen gleitenden Übergang von Homo erectus zu Homo sapiens annimmt. Im Sinne der Vermischungshypothese deutet Trinkaus beispielsweise ein 1998 im Lapedo-Tal in Zentralportugal gefundenes Kinderskelett („Kind von Lagar Velho“) als „Mischlingskind“. Das fast vollständig erhaltene Skelett des etwa vier Jahre alten Kindes war vor ca. 25.000 Jahren in einem Bett aus verbrannten Kiefernzweigen und bedeckt mit rotem Ocker bestattet worden, wie dies für die Beerdigungsrituale des anatomisch modernen Menschen und im Besonderen im Gravettien üblich war. Auch die meisten anatomischen Merkmale weisen es als modernen Menschen aus. So gleicht beispielsweise die Lage der Bogengänge des Innenohrs der des modernen Menschen und nicht des Neandertalers. Das Kinderskelett besitzt allerdings drei Auffälligkeiten: sein nach hinten fliehender Unterkiefer, bestimmte Ansätze der Brustmuskulatur und relativ kurze Unterschenkel. Diese drei Merkmale können auch bei Neandertalern auftreten. Die Reaktionen auf die Interpretation des Fossils waren im wissenschaftlichen Bereich jedoch überwiegend abweisend.
In einer Publikation von Erik Trinkaus und rumänischen Kollegen aus dem Jahre 2006 wurde gleichfalls argumentiert, dass die 1952 in der Höhle von Peștera Muierii in Rumänien entdeckten, 30.000 Jahre alten Knochen auf Gemeinsamkeiten zwischen Neandertaler und modernem Menschen hinweisen. Für Homo sapiens charakteristisch sei die vergleichsweise kleine Kinnlade mit den ausgeprägten Eckzähnen sowie kleine Augenbrauenbögen und enge Nasenöffnungen. Für den Neandertaler charakteristisch sei der große Augenabstand und die fliehende Stirn mit großen Überaugenwülsten. Unterschiedliche Auffassungen gab es auch um die rumänischen Peștera-cu-Oase-Funde, insbesondere um den 42.000 bis 37.000 Jahre alten Unterkiefer Oase 1.
Die große Mehrheit der Paläoanthropologen lehnte diese Vermischungshypothese jedoch ab, da die Fossilien beider Arten anhand zahlreicher anatomischer Merkmale in aller Regel deutlich zu unterscheiden seien:
„Spuren eines solchen möglichen Intermezzos wären dann aber deutlich in der Anatomie nachzuweisen, wenn man bedenkt, dass der stämmige Neandertaler der Eiszeit trotzte und der grazilere Homo sapiens der Sonne Afrikas entflohen war. Doch sämtliche gefundenen Schädel- und Skelettteile wurden – wenn auch nach einigen Irrungen und Wirrungen – entweder als eindeutig modern oder eindeutig neandertaloid eingestuft. Erst die Entdeckung des sogenannten ‚Mischlingskindes‘ aus Lagar Velho 1998 in Portugal gab den Vertretern der Vermischungstheorie neuen Auftrieb.“
Zudem wurde auch aus den Analysen der Neandertaler-DNA abgeleitet, dass der 2010 berichtete Genfluss vermutlich bereits vor der Besiedelung Europas durch den anatomisch modernen Menschen erfolgte. Die von Trinkaus herausgestellten anatomischen Merkmale wurden daher von der überwiegenden Mehrzahl der Paläoanthropologen als Hinweis auf die gemeinsame Abstammung von Neandertaler und anatomisch modernem Menschen aus Homo erectus gedeutet und nicht als Resultat einer genetischen Vermischung. Diese Auffassung wird auch durch Methoden der geometrischen Morphometrie gestützt. In einer Studie wurde zunächst die Größe der morphologischen Unterschiede zwischen Neandertaler und Homo sapiens bestimmt; im nächsten Schritt wurden jeweils paarweise die Unterschiede zwischen 12 heute lebenden Primaten-Arten bestimmt und die Größe dieser Unterschiede mit jener des Paares Neandertaler / Homo sapiens verglichen. Als Ergebnis wurde festgestellt, dass die morphologische Distanz von Neandertaler und Homo sapiens vergleichbar ist mit den morphologischen Unterschieden zwischen den heute lebenden Primaten-Arten. Auch erwies sich der jahrzehntelang dem Neandertaler zugeordnete Unterkiefer von Mezzena als Homo sapiens zugehörig.
- Eine 2013 durchgeführte DNA-Analyse des rund 40.000 Jahre alten Unterkiefers Oase 1 aus Rumänien ergab dann aber, dass es sich zumindest bei dem jungen Mann, von dem dieser Kiefer stammte, tatsächlich um den Nachfahren eines Mischlings handelte: 6 bis 9 Prozent seines Genoms wurde als vom Neandertaler stammend interpretiert. Das Chromosom 12 enthielt sogar einen extrem langen, dem Neandertaler zugerechneten Abschnitt, der ungefähr die Hälfte der Basenpaare dieses Chromosoms umfasste. Hieraus wurde abgeleitet, dass die Paarung nicht länger als vier bis sechs Generationen zurück gelegen haben kann, da andernfalls – durch Crossing-over – Veränderungen der Basenpaar-Sequenz aufgetreten wären. Nachgewiesen wurde jedoch auch, dass die vom anatomisch modernen Menschen stammenden DNA-Abschnitte eher Fossilienfunden aus Asien ähneln und nicht den späteren Funden aus Europa oder den heute lebenden Menschen; hieraus wurde abgeleitet, dass die Population, zu der Oase 1 gehörte, eine „Sackgasse“ repräsentiere, dessen Gene in die heutige Population des Homo sapiens nicht eingingen.
- Bei einem weiteren, im Jahr 2015 veröffentlichten Fund eines frühen Homo sapiens, dem Oberschenkelknochen von Ust-Ischim in Sibirien, wurde ebenfalls Neandertaler-DNA festgestellt und zwar mit einem Anteil von 2 Prozent. Der Zeitpunkt des Genflusses wurde auf rund 7.000 bis 13.000 Jahre vor Lebzeiten des Individuums, das vor ca. 45.000 Jahren lebte, datiert – mit genetischer Nähe zu den in Eurasien lebenden Menschen.
Diese Studien belegen, dass Verpaarungen zwischen Neandertaler und Cro-Magnon-Mensch (bzw. frühem eurasischen Homo sapiens) nicht nur in der Levante, sondern auch im Osten Europas und in Sibirien stattgefunden haben.
Vermischung des Erbmaterials

Ab Mitte der 1990er-Jahre wurden in der Arbeitsgruppe von Svante Pääbo am Max-Planck-Institut für evolutionäre Anthropologie Methoden entwickelt, fossile DNA – sogenannte aDNA – aus Neandertaler-Knochen zu gewinnen. Das Ziel dieser Forschungsarbeiten wurde wie folgt angegeben: „Vergleiche des menschlichen Genoms mit dem Genom von Neandertaler und Menschenaffen ermöglichen es, charakteristische Merkmale zu identifizieren, die den anatomisch modernen Menschen von allen anderen homininen Arten abheben.“
Wie oft gemeinsame, fruchtbare Nachkommen von Neandertalern und anatomisch modernen Menschen gezeugt wurden, ist nicht bekannt. Aus einer Autorengruppe um Svante Pääbo, die 2014 die Häufigkeit von Neandertaler-Allelen in den Autosomen mit deren Häufigkeit in den X-Chromosomen der heute lebenden Menschen verglich, hieß es jedoch, dies sei möglicherweise nur viermal vorgekommen.
Erste Befunde
Die Untersuchung der Mitochondrien-DNA (mtDNA) von zahlreichen Neandertaler-Skeletten, frühen Homo-sapiens-Funden und der vermuteten Mosaikformen am Max-Planck-Institut für evolutionäre Anthropologie lieferte keine Anhaltspunkte dafür, dass Gene von Homo neanderthalensis in Populationen des Homo sapiens übergegangen waren. Bereits 1997 wurden 378 Basenpaare der mtDNA vom Typusexemplar Neandertal 1 mit einer großen Vergleichsserie (rezenter) moderner Menschen verglichen. Im Ergebnis wurde damals geschlussfolgert, dass der gemeinsame Vorfahre etwa 600.000 Jahre vor heute gelebt habe und später kein erkennbarer genetischer Austausch mehr stattgefunden habe.
Parallel zur Sequenzierung der Mitochondrien-DNA versuchte man am selben Institut, die DNA aus dem Zellkern von Neandertalern zu rekonstruieren. Im Mai 2006 berichteten die Forscher dann, dass sie aus einem in der Vindija-Höhle in Kroatien gefundenen, 45.000 Jahre alten, männlichen Neandertaler rund eine Million Basenpaare – von insgesamt mehr als drei Milliarden – sequenzieren konnten. Eines der Ergebnisse dieser vorläufigen Analysen war, dass sich das Y-Chromosom des Neandertalers stärker vom Y-Chromosom des modernen Menschen und von dem des Schimpansen unterscheidet als die anderen Chromosomen. Insgesamt wurde aus diesen DNA-Analysen – zumindest für die späten Neandertaler – abgeleitet, dass kaum Vermischung mit anatomisch modernen Menschen stattgefunden habe.No Sex with Homo sapiens („Kein Geschlechtsverkehr mit Homo sapiens“) hieß es folgerichtig noch 2009 in einer Schrift des Max-Planck-Instituts für evolutionäre Anthropologie unter Verweis auf deren Experten für die DNA-Sequenzierung des Neandertaler-Genoms.
Für Aufsehen sorgte 2007 eine Detailanalyse: Der Bau des Gens MC1R und des von ihm codierten Rezeptorproteins MC1R ergab Hinweise darauf, dass möglicherweise einzelne Neandertaler eine sehr geringe Hautpigmentierung aufgewiesen haben könnten, vergleichbar der Pigmentierung von heute lebenden rothaarigen und zugleich hellhäutigen Menschen. MC1R reguliert nicht nur beim Menschen, sondern auch bei vielen anderen Wirbeltieren die Färbung der Haut.
Publikation des Genoms
Seit 2009 wurden erste Teilergebnisse der Entschlüsselung des Genoms mitgeteilt. Die Genome des modernen Menschen und des Neandertalers unterscheiden sich um weniger als 0,5 Prozent, hieß es darin. Im Mai 2010 veröffentlichte ein Team um Richard E. Green aus der Arbeitsgruppe von Svante Pääbo schließlich eine ausführliche Analyse der rekonstruierten 60 Prozent aDNA des Neandertalers (die Seitenzahlen im folgenden Text beziehen sich auf diese Studie). Eine noch umfassendere Rekonstruktion der DNA wurde schließlich im Frühjahr 2013 publiziert.
Ausgangsmaterial waren aDNA-Fragmente von weniger als 200, zumeist nur 40 bis 60 Basenpaaren. Sie waren drei Bruchstücken von Schienbein-Funden aus der Vindija-Höhle entnommen und ihre Rekonstruktion ergänzend mit aDNA aus drei weiteren Neandertalerfunden verglichen worden, von denen einer der Holotypus aus dem Neandertal war. Die rekonstruierte Neandertaler-DNA wurde anschließend mit DNA-Proben moderner Menschen aus Afrika, Europa und Asien verglichen.
Die DNA-Analyse ergab erneut, dass die Erbanlagen der Neandertaler sich nur in sehr geringem Maße von den Erbanlagen der heutigen Menschen unterscheiden. Die Autoren der Studie betonten sogar ausdrücklich „die Tatsache, dass die Neandertaler innerhalb der Variationsbreite der modernen Menschen liegen“ (S. 713). Gleichwohl wurden mehrere Dutzend Genvarianten identifiziert, anhand derer Neandertaler und Homo sapiens unterschieden werden können. Von einigen gilt als gesichert, dass sich die verursachenden Mutationen erst nach der Trennung der beiden Entwicklungslinien ereigneten, die zu Neandertalern und anatomisch modernen Menschen führten: Und zwar beim anatomisch modernen Menschen, denn diese Gene haben beim Neandertaler und bei den heute lebenden Schimpansen eine identische – das heißt ursprüngliche – Nukleotidsequenz. Herausgestellt wurden unter anderem Veränderungen am Gen SPAG17, das den Bau der Axonemen und hierdurch die Beweglichkeit der Geißel eines Spermiums beeinflusst, ferner am Gen PCD16, das in Fibroblasten für Cadherin-1 codiert und möglicherweise die Wundheilung beeinflusst. Drei weitere Gene beeinflussen Struktur und Pigmentierung der Haut (S. 715).
Andere Mutationen betreffen Gene, bei denen ein Zusammenhang mit kognitiven Funktionen des Menschen besteht; Abweichungen von der „genetischen Norm“ machen sich bei den betroffenen Menschen in Form einer erhöhten Auftretenswahrscheinlichkeit bestimmter psychischer Störungen bemerkbar.
Herausgestellt wurde auch das im Vergleich zum Neandertaler beim anatomisch modernen Menschen veränderte Gen RUNX2, bei dem eine Fehlfunktion Abweichungen im Bau des Kopfes und des Rumpfes zur Folge hat. Die Autoren der Studie vermuteten, „dass diese Veränderung Merkmale der Morphologie des Oberkörpers und des Kopfes beeinflusste“ (S. 717). Anhaltspunkte dafür, dass bestimmte äußerlich erkennbare Merkmale des anatomisch modernen Menschen durch Neandertaler-Gene verursacht werden, wurden nicht entdeckt.
Nachweis von Genfluss zum Neandertaler
Forscher des Max-Planck-Instituts für evolutionäre Anthropologie konnten 2016 in Funden einer Höhle im sibirischen Altai-Gebirge Gene aus dem Zellkern des modernen Menschen nachweisen. Sie kommen zu dem Schluss, dass die Vermischung vor rund 100.000 Jahren stattgefunden hat. Die Vermischung liegt damit deutlich früher als die bisher in Europa nachweisbare, die auf circa 60.000 Jahre angesetzt wird. Moderne Menschen und Neandertaler trafen nach Ansicht der Forscher wahrscheinlich zuerst im Nahen Osten aufeinander und dieser Vorgang wiederholte sich zu anderen Zeiten an anderen Stellen.
Eine Untersuchung der Mitochondrien-DNA (mtDNA) aus dem Jahr 2017 erbrachte Hinweise auf einen weiteren Genfluss in der Zeit zwischen 470.000 und 220.000 Jahren vor heute vom anatomisch modernen Menschen (Homo sapiens) hin zum Neandertaler. Es zeigte sich ein fundamentaler Unterschied in der mtDNA früher und später Neandertaler-Funde: Die mtDNA früher Fossilien ist ähnlich der des Denisova-Menschen, während die mtDNA später Neandertaler der des anatomisch modernen Menschen ähnelt. Die Ergebnisse passen zur Hypothese, dass sich zwischen 765.000 und 550.000 Jahren vor heute die Entwicklungslinien zum Neandertaler und Denisova-Menschen einerseits und zum anatomisch modernen Menschen andererseits aufspalteten. Danach kam es zur Trennung der Neandertaler- und Denisova-Linie, was die Denisova-ähnliche mtDNA der frühen Neandertaler erklärt. Diese wurde später – so die Annahme – durch mtDNA der modernen Menschen ersetzt, so dass junge Neandertaler-Funde eine im Vergleich zum Denisova-Menschen weitaus ,modernere’ mtDNA aufweisen. Anatomisch moderne Menschen wären also schon relativ früh von Afrika nach Europa gewandert, ohne dort zu überdauern, oder diese „Vorhut“ wäre in den damals weit verbreiteten Neandertaler-Populationen aufgegangen.
Die genetische Vielfalt und damit auch die Verbreitung der Neandertaler erscheint größer als vor der Untersuchung gedacht. Erst in der Neandertaler-Spätzeit waren dessen Populationen so ausgedünnt, dass er dem erneuten Migrationsdruck des Homo sapiens weichen musste. Für diese Untersuchungen wurden neben den Denisova-Funden und 430.000 Jahre alten Frühmenschenfossilien aus der Höhle Sima de los Huesos in Nordspanien (mit der Denisova-ähnlichen mtDNA) ein etwa 124.000 Jahre alter Neandertaler-Oberschenkelknochen von der Schwäbischen Alb, der 1937 in der Hohlenstein-Stadel-Höhle nahe Ulm gefunden wurde (und dessen mtDNA die ,modernen’ Merkmale trug) herangezogen. Der Oberschenkelknochen aus der Höhle bei Ulm ist das derzeit älteste Neandertaler-Fossil, dessen mtDNA die modernen Merkmale zeigt und grenzt daher den Zeitraum für die genetische Infiltration zusammen mit dem nordspanischen Fund ein.
Die Annahme liefert auch eine Erklärung für das Paradoxon, dass nach Genanalysen der Zellkern-DNA (nukleären DNA) der nächste Verwandte des Neandertalers der Denisova-Mensch ist; nach den Analysen der mtDNA (zunächst nur von späten Neandertaler-Fossilien) schien dies aber der moderne Mensch zu sein. – Für die alten Fossilien aus Sima de los Huesos zeigt die mtDNA dagegen die gleiche Verwandtschaft an wie die Zellkern-DNA.
Im Jahr 2020 wurden Hinweise darauf publiziert, dass es vor maximal 370.000 Jahren, spätestens aber vor 100.000 Jahren, zu einem Übergang des Y-Chromosoms von Homo sapiens in die Population der Neandertaler gekommen sein könnte.
Nachweis von Genfluss zu Homo sapiens

Besonderes Augenmerk richteten die Forscher bereits im Jahr 2010 auf den Vergleich des Neandertaler-Genoms mit menschlicher DNA aus unterschiedlichen Erdteilen und Ethnien: mit DNA-Sequenzen eines Franzosen, eines Han-Chinesen, eines Papua, eines Yoruba und eines San. Sie berichteten, dass das Genom der Neandertaler eine signifikant größere Ähnlichkeit mit dem Genom von Europäern und Asiaten hat als mit dem Genom von Afrikanern: Der Franzose, der Han und der Papua stehen den Neandertalern in gleichem Maße nahe, der Yoruba und der San weisen diese genetische Nähe gleichermaßen nicht auf (S. 718). Die Autoren deuteten dies so: „Die sparsamste Erklärung für diese Beobachtung ist, dass Neandertaler Gene mit den Vorfahren der Nichtafrikaner austauschten.“ Da anhand weiterer Analysen der untersuchten Genome der fünf Vertreter heutiger Populationen ein Genfluss vom Homo sapiens zum Neandertaler ausgeschlossen werden konnte, kam die Studie zu dem Ergebnis „dass der Genfluss vom Neandertaler zu den Vorfahren der Nichtafrikaner erfolgte, bevor sich die eurasischen Gruppen voneinander trennten“ (S. 710), das heißt im Nahen Osten, wo Neandertaler und anatomisch moderne Menschen in der Zeitspanne von vor 110.000 Jahren bis vor rund 50.000 Jahren koexistierten.
Gestützt wird diese Vermutung, der Genfluss sei ausschließlich in eine Richtung gegangen, unter anderem durch eine Studie, der zufolge die Wahrscheinlichkeit grundsätzlich sehr viel größer ist, dass Gene von einer ortsansässigen Population in eine andere Population übergehen, wenn diese andere Population in das Siedlungsgebiet der ansässigen Population eindringt, als umgekehrt. 2012 wurde die Zeitspanne des möglichen Genflusses in die Zeit vor 65.000 bis 47.000 Jahren eingegrenzt.
Das Ausmaß des Genflusses vom Neandertaler zu Homo sapiens beträgt den Autoren der Studie zufolge zwischen einem und vier Prozent des Genoms der heutigen nichtafrikanischen Bevölkerung (S. 721). Sie äußerten zugleich ihr Erstaunen darüber, dass der Genfluss in Europa nicht größer war als in Asien, obwohl in Europa einige Fossilien gefunden worden seien, die von Vertretern der „Vermischungshypothese“ als Beleg für einen solchen Genfluss angeführt wurden. Sie schlossen nicht aus, dass spätere Wanderungsbewegungen im Zusammenhang mit der Verbreitung der Landwirtschaft früheren Genfluss verdeckt haben. Möglich sei aber auch, dass – im Unterschied zur anfänglichen Situation im Nahen Osten – in Europa bereits relativ große Populationen von Homo sapiens auf die Neandertaler trafen; ein in geringem Maße erfolgter Genfluss sei dann heute nicht mehr nachweisbar. Die „Kreuzungsrate“ – bezogen auf die Anzahl später eigenen Nachwuchs zeugender Neugeborener – betrug Modellrechnungen zufolge weniger als zwei Prozent, vermutlich sogar weniger als ein Prozent.
2011 interpretierte eine kanadische Forschergruppe die Übereinstimmungen eines Abschnitts des X-Chromosoms in Populationen außerhalb Afrikas mit jenem des Neandertalers bei gleichzeitigem Fehlen solcher Übereinstimmungen in afrikanischen Populationen als weiteren Beleg für einen Genfluss vom Neandertaler zu Homo sapiens.
Dieser Deutung wurde jedoch 2012 anhand einer Modellrechnung widersprochen und eine andere Interpretation der genetischen Befunde vorgelegt: Die größere Übereinstimmung des Genoms der außerafrikanischen Populationen von Homo sapiens mit dem Genom der Neandertaler könne auch dadurch erklärt werden, dass zufälligerweise eine Population des Homo sapiens Afrika verlassen habe, die noch eine besonders große genetische Ähnlichkeit mit dem gemeinsamen Vorfahren der anatomisch modernen Menschen und der Neandertaler hatte. Eine andere Modellrechnung kam allerdings zu dem umgekehrten Ergebnis: Deren Autoren argumentierten, es sei sehr unwahrscheinlich, dass archaische Bevölkerungsstrukturen in Afrika die genetischen Übereinstimmungen zwischen Neandertalern und anatomisch modernen Menschen außerhalb von Afrika erklären könnten.
Anfang 2014 wurden weitere, verfeinerte Analysen des Genflusses von Neandertalern zu Homo sapiens publiziert. Laut einer Veröffentlichung von Autoren der Harvard Medical School beträgt der Anteil des Neandertaler-Genoms in den Autosomen der heute lebenden Europäer 1,15 % und in denen der Ostasiaten 1,38 %, in den X-Chromosomen beider Bevölkerungsgruppen hingegen nur rund 0,20 bis 0,30 %, also nur rund ein Fünftel des Anteils in den Autosomen. Daraus wurde geschlossen, dass – vergleichbar mit anderen Säugetieren – die Fruchtbarkeit der männlichen Mischlinge reduziert war. Besonders häufig wurden von Neandertalern abstammende Allele in Regionen des Genoms heute lebender Menschen nachgewiesen, die mit der Codierung von Keratinfasern in Zusammenhang gebracht werden; möglicherweise – so wurde spekuliert – haben diese Allele den anatomisch modernen Menschen die Anpassung an den außerafrikanischen Lebensraum erleichtert. Gestützt wurden diese Befunde durch die Studie einer zweiten, unabhängigen Forschergruppe der University of Washington; auch deren Analysen erbrachten Hinweise auf eine reduzierte Fitness der Mischlinge. Zusätzlich wurde in dieser Studie der Einfluss von Neandertaler-Allelen auf die Bildung von Keratinen erläutert: Demnach ist das Gen BNC2 auf Chromosom 9 an der Pigmentierung der Haut beteiligt und ist insbesondere mit der (relativ hellen) Pigmentierung der Europäer verbunden. Schließlich wurde in einer weiteren unabhängigen Studie nachgewiesen, dass das Y-Chromosom des (männlichen) Neandertalers ein mutiertes Gen aufweist, das bei Schwangeren zu einer Immunantwort führen kann, in deren Folge die Wahrscheinlichkeit einer Fehlgeburt erhöht ist. 2021 wurde berichtet, dass lediglich 1,5 bis 7 % des modernen menschlichen Genoms einzigartig ist, den Rest der Allele findet man auch beim Neandertaler und/oder beim Denisovaner.
Siehe auch
Literatur
- Historische Forschung
- C. Fuhlrott: Menschliche Ueberreste aus einer Felsengrotte des Düsselthals. Ein Beitrag zur Frage über die Existenz fossiler Menschen in der Google-Buchsuche. In: Verhandlungen des naturhistorischen Vereins der preussischen Rheinlande und Westphalens. Band 16, 1859, S. 131–153.
- Gustav Schwalbe: Der Neanderthalschädel. In: Bonner Jahrbücher, 106, A. Marcus und E. Weber, Bonn 1901, S. 1–71, Tafel I. (Archive)
- Gustav Schwalbe: Die Vorgeschichte der Menschen. Friedrich Vieweg und Sohn, Braunschweig 1904. (Archive)
- Gustav Schwalbe: Studien zur Vorgeschichte des Menschen, E. Schweizerbart’sche Verlagsbuchhandlung (E. Nägele), Stuttgart 1906. (Archive)
- Aktuelle Forschung
- Paul Mellars: Neanderthals and the modern human colonization of Europe. Review-Artikel in: Nature. Band 432, 2004, S. 461–465, doi:10.1038/nature03103, Volltext (PDF).
- Nicholas J. Conard, Jürgen Richter: Neanderthal Lifeways, Subsistence and Technology. One Hundred and Fifty Years of Neanderthal Studies. Springer 2011, ISBN 978-94-007-0415-2.
- Kay Prüfer, Fernando Racimo et al.: The complete genome sequence of a Neanderthal from the Altai Mountains. In: Nature. Band 505, Nr. 7481, 2014, S. 43–49, doi:10.1038/nature12886, Volltext (PDF).
- Wil Roebroeks und Marie Soressi: Neandertals revised. In: PNAS. Band 113, Nr. 23, 2016, S. 6372–6379, doi:10.1073/pnas.1521269113. (freier Volltext)
- Kay Prüfer et al.: A high-coverage Neandertal genome from Vindija Cave in Croatia. In: Science. Band 358, Nr. 6363, 2017, S. 655–658, doi:10.1126/science.aao1887
- Michael Dannemann und Janet Kelso: The Contribution of Neanderthals to Phenotypic Variation in Modern Humans. In: The American Journal of Human Genetics. Band 101, Nr. 4, 2017, S. 578–589, doi:10.1016/j.ajhg.2017.09.010, Volltext.
- Patrick F. Reilly et al.: The contribution of Neanderthal introgression to modern human traits. Review-Artikel in: Current Biology. Band 32, Nr. 18, 2022, PR970–R983, doi:10.1016/j.cub.2022.08.027. (freier Volltext)
Filme
- Am Anfang war das Feuer. Spielfilm, Frankreich 1981, Regie: Jean-Jacques Annaud.
- Ao, der letzte Neandertaler. Spielfilm, Sunfilm Entertainment, Frankreich 2010, Regie: Jacques Malaterre.
- Einer von uns: Der Homo sapiens. Teil 4. Europa: Homo sapiens et neanderthalensis. Dokumentarfilm, Arte 2015. (Video auf YouTube, Inhaltsübersicht.)
- Auf den Spuren der Neandertaler - Das Rätsel der Bruniquel-Höhle. Dokumentarfilm von Luc-Henri Fage. Arte, 2019 (53:26 min).
- Schwierige Verwandtschaft: Die Neanderthaler und wir. Gerd-Christian Weniger, Direktor des Neanderthal Museums Mettmann und Professor am Institut für Ur- und Frühgeschichte der Universität zu Köln. Vortrag vom 8. November 2012 auf YouTube.
- Das Neandertaler Rätsel. Brudermord in der Steinzeit? Von Thomas Cirotteau. ZDFinfo, 10. August 2020, 44 min (Video auf YouTube).
Weblinks
- Museen
- LVR LandesMuseum Bonn: Ausstellungsort der originalen Fossilien des Neandertalers aus dem Neandertal.
- Neanderthal-Museum im Neandertal: Website des Neanderthal Museums unweit der Fundstelle des Neandertalers aus dem Neandertal. Die Besucher durchmessen im Museum eine Rekonstruktion der Evolution des Menschen.
- Krapina Neanderthal Museum. Beschreibung des Neandertalermuseums in Krapina, Kroatien (englisch)
- Populäre Darstellungen
- Urzeit: Neandertaler. Auf: planet-wissen.de, einem Medienangebot des WDR.
- Die Neandertaler auf der Website der Max-Planck-Gesellschaft.
- Neandertaler so kunstsinnig und intelligent wie wir? Auf: spektrum.de vom 11. Juni 2013.
- Sheanderthal. Not all Neanderthals were ‘cavemen’: half were women. Essay der britischen Archäologin Rebecca Wragg Sykes.
- Wissenschaftliche Darstellungen
- The Neandertal Genome. (Memento vom 7. März 2015 im Internet Archive). Erschienen auf sciencemag.org im Herbst 2010.