
| Robben | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Ein pazifisches Walross | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pinnipedia | ||||||||||||
| Illiger, 1811 | ||||||||||||
| Familien | ||||||||||||
|
Die Robben (Pinnipedia) sind ein Taxon im Wasser lebender Raubtiere (Carnivora) und gehören somit ökologisch zu den Meeressäugern. Die wissenschaftliche Bezeichnung „Pinnipedia“, abgeleitet von den lateinischen Wörtern pinna ‚Flosse‘ und pes ‚Fuß‘, bedeutet Flossenfüßer. Sowohl diese Bezeichnung als auch die Bezeichnung Wasserraubtiere findet man mitunter auch in der Fachliteratur. In einer älteren biologischen Systematik wurden die Wasserraubtiere den nicht mehr als Taxon anerkannten „Landraubtieren“ (Fissipedia) gegenübergestellt.
Anatomie

Robben sind mittelgroße bis große Säugetiere, die Längen zwischen 1,20 und 6 Metern erreichen können. Das Gewicht schwankt zwischen den Arten extrem und liegt zwischen 25 Kilogramm bei Weibchen der Antarktischen Seebären (Arctocephalus gazella) und mehr als 4 Tonnen bei männlichen Südlichen See-Elefanten (Mirounga leonina). Oft lässt sich ein auffälliger Sexualdimorphismus feststellen: See-Elefanten-Männchen wiegen oft bis zu viermal mehr als ihre Weibchen. Es gibt jedoch auch einige Arten, bei denen das Weibchen größer ist, z. B. den Seeleopard.
Der Robbenkörper hat eine torpedoförmige Gestalt: Der abgeflachte Kopf ist durch einen dicken Hals nur undeutlich vom Rest des Körpers abgesetzt, der Schwanz nur noch als Rudiment erhalten. Die Gliedmaßen sind zudem bis zum Ellenbogen beziehungsweise Knie in den Körper eingebettet, so dass nur Unterarm und -schenkel frei liegen. Alle diese Eigenschaften setzen den Strömungswiderstand herab und dienen so der Anpassung an das Leben im Wasser.
Schädel und Gebiss
Im Robbenschädel überlappt das vordere Hinterhauptsbein, der Supraokzipitalknochen mit dem Schläfenbein, dem Parietalknochen; die Augenhöhlen sind vergrößert, während das Rostrum, also die „Schnauze“ des Tieres, stark verkürzt ist. Statt des typischen Raubtiergebisses haben die meisten Robben ein eher gleichförmiges Fischfressergebiss. Es existieren meist ein bis zwei Paar untere Schneidezähne, kegelförmige, meist wenig betonte Eckzähne und zwölf bis vierundzwanzig einfache, homodonte, also gleichartig aufgebaute Backenzähne. Letztere besitzen zwei Wurzeln und sind zugespitzt; sie sind somit weniger zum Kauen als zum Festhalten der Beute geeignet. Reißzähne wie bei den anderen Raubtieren kommen bei Robben nicht vor. Allerdings gibt es zahlreiche Abweichungen, vor allem bei jenen Robbenarten, die andere Ernährungsweisen pflegen: Bei Walrossen sind die Eckzähne etwa als Stoßzähne ausgebildet, während bei Krabbenfressern die Zähne kompliziert gebaute Höcker besitzen, die bei geschlossenem Kiefer ein engmaschiges Sieb bilden, mit dem die Tiere ihre Nahrung, Krill, erbeuten.
Wirbelsäule und Gliedmaßen

Je nach Familie sind unterschiedliche Teile der Wirbelsäule verstärkt, dies hängt mit der unterschiedlichen Fortbewegung der Tiere zusammen. Bei Ohrenrobben, bei denen die Vorderflossen im Mittelpunkt stehen, sind die Hals- und Brustwirbel deutlich verstärkt, während bei Hundsrobben, die sich mithilfe ihrer Hinterflossen fortbewegen, stattdessen die Lendenwirbel vergrößert sind. Zusätzlich sind bei ihnen die Gelenkfortsätze der Wirbel stark reduziert, so dass diese nicht so starr miteinander verbunden sind wie bei anderen Säugetieren. Durch diese Flexibilität kommt die große Wendigkeit des Robbenkörpers zustande. Diese wirkt sich durch das fehlende Schlüsselbein (Clavicula) und den kleinen, parallel zur Wirbelsäule ausgerichteten Beckengürtel auch auf die Bewegungsfreiheit der Gliedmaßen aus.
Alle vier Beine wurden im Laufe der Evolution zu Flossen umgewandelt. Oberarmknochen (Humerus), Elle (Ulna) und Speiche (Radius) sind im Vergleich zu anderen Raubtieren kürzer, aber kräftiger ausgeprägt, auch der Oberschenkelknochen (Femur) ist flach und breit, so dass eine effektive Kraftübertragung im Wasser ermöglicht wird. Jede der Flossen endet in fünf langen, abgeflachten Zehen, die durch Schwimmhäute miteinander verbunden sind; oft sind die erste und die fünfte Zehe verlängert. Eine weitere Spezialisierung der Flossen wurde vermutlich nur durch die fortbestehende Notwendigkeit, sich an Land fortzubewegen, verhindert.
Fell und Fettschicht
Geboren werden Robben meistens mit einem dichten Fell, das beim Älterwerden verschwindet und durch ein kurzes Haarkleid ersetzt wird. Nur die Seebären behalten auch als Alttiere ein auffälliges Pelzkleid, das aus einem dichten Unterfell und darüber herausragenden steifen Grannenhaaren besteht; Walrosse haben ein kurzes, wenig auffälliges Haarkleid. Bei Ohrenrobben wird das Fell zusammen mit der obersten Hautschicht über einen längeren Zeitraum hinweg, der bis zu einem Monat betragen kann, erneuert, während Hundsrobben diesen Häutungsprozess meist verhältnismäßig abrupt innerhalb weniger Tage vollziehen.
Zum Schutz vor Auskühlung befindet sich unterhalb der Haut eine bei manchen Arten bis zu zehn Zentimeter dicke Fettschicht. Diese Schicht dient daneben als Nährstoffspeicher und trägt im Wasser zum Auftrieb bei. Sie ist bei Hundsrobben und Walrossen besonders ausgeprägt. Allerdings schützt sie nicht die durch eine besonders große Zahl von Blutgefäßen versorgten Flossen.
Innere Organe
Der Robbenmagen ist ein einfacher länglicher Schlauch und darauf eingerichtet, als Ganzes verschluckte Beute aufzunehmen. Sehr aufwendig und lang ist dagegen der Dünndarm, der bei Männchen der Südlichen See-Elefanten (Mirounga leonina) eine Länge von über 200 Metern erreichen kann; alle anderen Darmabschnitte sind dagegen verhältnismäßig kurz.
Nicht mehr äußerlich sichtbar, weil weitgehend ins Körperinnere verlagert, sind die Hoden der Männchen; eine Ausnahme stellen lediglich die Walrosse dar, bei denen die Geschlechtsorgane besonders in der Paarungszeit gut zu erkennen sind. Auch die Zitzen der Weibchen liegen nicht mehr auf der Oberfläche: Da Jungtiere die Zitzen so nicht mehr mit dem Maul umfassen können, muss das Muttertier die Milch bei Bedarf aktiv ins Maul seines Jungen spritzen.
Sinnesorgane
Die Sinnesorgane der Robben müssen sowohl unter Wasser als auch an Land funktionieren und sind daher einander widerstrebenden Anforderungen ausgesetzt.
Augen
Die Augen sind verhältnismäßig groß und ermöglichen eine gute Unterwassersicht. An die oft geringe Lichtintensität unter Wasser sind sie durch eine stark vergrößerte Anzahl an Stäbchen in der Netzhaut angepasst, die im Gegensatz zu den Zapfen zwar keine Farbsicht ermöglichen, dafür aber wesentlich lichtempfindlicher sind. Das Empfindlichkeitsspektrum variiert mit dem Lebensraum: Das Maximum liegt bei tieftauchenden Arten wie zum Beispiel See-Elefanten (Mirounga) im blauen Bereich, ist aber etwa bei Seehunden (Phoca vitulina), die sich vornehmlich in Küstengewässern aufhalten, in Richtung Grün verschoben. Farbsicht ist bei Robben durch die eingeschränkte Zapfenanzahl nur in eingeschränktem Ausmaß möglich. Eine weitere Anpassung an die geringen Lichtverhältnisse ist das Tapetum lucidum, eine hinter der Netzhaut gelegene reflektierende Schicht, die Licht, das die Netzhaut durchquert hat, zurückwirft: So besteht die Möglichkeit, Licht, das beim ersten Passieren nicht wahrgenommen wurde, beim zweiten Mal zu registrieren.
Dem Leben im Wasser ist nicht nur eine oft geringe Lichtintensität zu eigen, sondern auch eine andere Lichtbrechung: So hat die Hornhaut nahezu den gleichen Brechungsindex wie Wasser und fällt daher unter Wasser als Bestandteil des Abbildungsapparats aus. Als Ausgleich ist bei Robben die Augenlinse nahezu kugelförmig ausgebildet und besitzt daher eine wesentlich höhere Brechkraft. Weil in Luft Linse und Hornhaut lichtbrechend wirken, sind Robben hingegen an Land, wo der Sehsinn eine geringere Bedeutung hat, stark kurzsichtig. Dafür sind sie in der Lage, die hohen Lichtintensitäten, die etwa von sonnenbeschienenen Eisflächen ausgehen, zu tolerieren, indem sie die Pupille zu einem engen senkrechten Schlitz verengen. Die Hornhaut bildet den äußeren Abschluss des Auges. Sie wird beständig mit Tränenflüssigkeit benetzt, um Fremdkörper abzuwaschen und den direkten Augenkontakt mit Salzwasser zu verhindern. Anders als ihre nahen Verwandten besitzen Robben jedoch keine Tränenkanäle.
Obwohl der Sehsinn auch für Robben eine große Bedeutung einnimmt, ist er nicht unersetzlich: Bei Seelöwen und Seebären, deren Männchen oft blutige Revierkämpfe durchführen, kommt es nicht selten zum Verlust des Augenlichts; die Tiere sind durch ihre Blindheit aber anscheinend wenig eingeschränkt und kommen relativ problemarm zurecht.
Gehör
Äußerlich sind die Ohren der Robben verkümmert oder nicht mehr existent; falls vorhanden, werden sie beim Tauchen verschlossen. Dennoch besitzen Robben ein exzellentes Gehör. Das Innenohr steht bei ihnen nur mit je einem einzigen Schädelknochen in Verbindung, wodurch die unterschiedslose Übertragung von Schallwellen durch den ganzen Schädel unterbunden wird; erst dadurch wird auch unter Wasser Richtungshören möglich. Die Auskleidung des Hörkanals und des Mittelohrs erlaubt beim Tauchen die Anpassung des Innendrucks.
Der wahrgenommene Frequenzbereich unterscheidet sich je nach Medium: Im Wasser können Robben wesentlich höhere Töne wahrnehmen als der Mensch; dafür ist das Gehör an Land bei etwa gleichem Wahrnehmungsspektrum weniger empfindlich.
Geruchs- und Tastsinn
Der Geruchssinn ist wie bei den meisten Landraubtieren sehr gut entwickelt; da die schlitzartigen Nasenöffnungen unter Wasser geschlossen sind, funktioniert er aber nur an Land. Er spielt insbesondere bei Begegnungen mit Artgenossen eine Rolle: So wird die Mutter-Kind-Beziehung oft durch den Geruchssinn aufrechterhalten; bei vielen Ohrenrobben dient er den Männchen dazu, die Empfängnisbereitschaft der Weibchen zu ermitteln.
Für die Orientierung im Wasser ist ein weiterer Sinn bedeutsam: der hoch entwickelte Tastsinn der in horizontalen Längsreihen auf der „Schnauze“ angeordneten Barthaare (Vibrissen). Jedes dieser Haare entspringt in einer reich mit Nervenendigungen und Blutgefäßen versorgten Bindegewebekapsel. Vibrissae können Druck- und Strömungsänderungen erspüren und dienen den Tieren vermutlich auch zum Abschätzen ihrer Geschwindigkeit. Zudem gibt es Hinweise darauf, dass die Tasthaare besonders empfindlich gegenüber niederfrequenten Schwingungen sind, die von der Bewegung von Beutetieren herrühren. Tatsächlich können Robben, die ihre Tasthaare verloren haben, selbständig keine Fische mehr fangen und müssen daher verhungern. Schließlich werden Vibrissae zur Kommunikation etwa bei Rangordnungskonflikten eingesetzt – aufgestellte Tasthaare signalisieren häufig die Bereitschaft zur Aggression.
Thermoregulation
Wie alle Säugetiere sind Robben endotherm, das heißt, ihr Körper weist eine nahezu konstante Innentemperatur auf. Um eine Auskühlung im Wasser und zugleich eine Überhitzung an Land zu vermeiden, haben die Tiere zahlreiche Anpassungen entwickelt.
Bereits die Körpergröße, die jene der landlebenden Raubtiere meist um ein Vielfaches übertrifft, verringert durch das wesentlich günstigere Verhältnis von wärmeproduzierendem Körpervolumen zu wärmeabgebender Körperoberfläche eine Auskühlung (siehe A/V-Verhältnis) und kann als Adaptation an das Medium Wasser gewertet werden. Die bereits erwähnte Fettschicht direkt unter der Haut ist wegen ihrer geringeren Wärmeleitfähigkeit zudem ein guter, von der Wassertiefe unabhängiger Isolator. Daneben tritt bei den Seebären das wasserdichte Fell, das vor allem durch darin gelöste Luftbläschen wärmerückhaltend wirkt. Die isolierende Luftschicht wird allerdings durch den mit der Wassertiefe zunehmenden Druck komprimiert und verliert daher bei tiefen Tauchgängen ihre Wirksamkeit.
Robben sind auch in der Lage, den Blutfluss in ihre Gliedmaßen zu regulieren. So können sie die Energieabgabe über die Flossen unter kalten Bedingungen so minimieren, dass die Temperatur knapp oberhalb der Gefriertemperatur bleibt. Spezielle Querverbindungen zwischen arteriellen und venösen Gefäßen, die arteriovenösen Anastomosen (AVA), erlauben umgekehrt in warmer Umgebung einen verstärkten Blutfluss in oberflächennahen Hautschichten. Bei Seebären sind sie nur in den Flossen vorhanden und der Wärmeverlust über diese Gliedmaßen ist somit die einzige Abkühlungsmöglichkeit. An Land schlagen die Tiere daher oft zusätzlich mit den Flossen, um den Energieaustausch mit der Umgebung durch den erzeugten Luftstrom zu maximieren. Eine weitere Möglichkeit besteht darin, die Flossen mit Urin zu benetzen, um dem Körper so durch die Verdunstungskälte Energie zu entziehen.
Sowohl Hundsrobben als auch Walrosse besitzen dagegen über die ganze Körperoberfläche hinweg arteriovenöse Anastomosen, so dass sie die Blutzirkulation und damit die Wärmeabgabe oder -aufnahme gezielt regulieren können. Auf diese Weise sind die Tiere in der Lage, sich selbst bei niedrigen Lufttemperaturen durch die bei der Absorption von Sonnenstrahlung freiwerdende Energie aufzuwärmen. Bei sonnenbadenden Walrossen lässt sich dieser Effekt wegen der fehlenden Behaarung anhand der rosaroten Hautfärbung besonders deutlich erkennen.
Daneben greifen manche Robben auch auf verhaltensbasierte Thermoregulation zurück: Bei drohender Überhitzung begeben sich etwa Mönchsrobben (Monachus) zurück ins Wasser. Dazu zählt auch die Tatsache, dass die energieintensive Häutung, die mit vermindertem Auskühlungsschutz verbunden ist, meist an Land stattfindet, wobei sich oft zahlreiche Tiere hautnah aneinanderlegen, um die Wärmeabgabe an die Umgebung zu vermindern.
Fortbewegung im Wasser und Anpassungen an das Tauchen
Im Wasser spielen Robben die Vorteile ihres Körperbaus aus. Der Vortrieb wird dabei bei den Ohrenrobben von den Vorder- und bei Walrossen und Hundsrobben von den Hinterflossen erzeugt. Sie haben dabei einige der energieeffizientesten Fortbewegungsmethoden des Tierreichs entwickelt.
Obwohl Robben zum Luftholen an die Oberfläche kommen müssen, können sie eine beträchtliche Zeit unter Wasser bleiben. Beim Tauchen kollabieren die Lungen, die darin enthaltene Luft wird durch Mund oder Nase ausgestoßen. Auf diese Weise befindet sich beim Tauchen kaum freies Gas im Robbenkörper und kann daher auch nicht wie bei menschlichen Tauchern beim Auftauchen zur Bildung von Blasen im Blut führen. Der gesamte während des Tauchvorgangs verfügbare Sauerstoff muss also in gebundener Form vorliegen. Das Blutvolumen und die Gesamtzahl roter Blutkörperchen ist bei Robben ebenso erhöht wie deren Gehalt an Hämoglobin, was zu einer deutlich gesteigerten Sauerstoffspeicherkapazität im Vergleich zu rein terrestrischen Säugetieren führt. Das Muskelgewebe der Tiere ist zudem gegenüber Landsäugetieren deutlich mit dem sauerstoffspeichernden Protein Myoglobin angereichert und toleriert große Mengen an Kohlenstoffdioxid (CO2).
Daneben verlangsamt sich der Herzschlag der Robben beim Tauchen erheblich. So kann sich die Herzaktivität von normalen hundert Schlägen pro Minute auf bis zu vier Schläge verringern, während gleichzeitig die Blutversorgung nicht unmittelbar lebensnotwendiger Organe eingeschränkt und die Körpertemperatur herabgesetzt wird. Diese Maßnahmen dienen der Verringerung des Sauerstoffverbrauchs während des Tauchens. Die Reduktion des Herzschlags und die Vasokonstriktion sind jedoch meist nicht so stark ausgeprägt, wie dies bei einem regulären Tauchreflex zu erwarten wäre. Darüber hinaus können Robben ihren Herzschlag in Erwartung der voraussichtlichen Tauchdauer regulieren.
Trotz diesen Anpassungen zur Sauerstoffspeicherung und Rationierung während des Tauchens, sind cerebrale Neuronen und Herzmuskelzellen der Robben regelmäßig längeren Phasen der Hypoxie ausgesetzt. Neuronen aus dem Gehirn der Klappmütze beispielsweise überdauern über zehn Minuten unter hypoxischen Bedingungen ohne langfristige Schädigungen, während Neuronen von Mäusen bereits nach fünf Minuten schwere Schädigungen davontragen können. Die grundlegenden Mechanismen dieser Hypoxietoleranz sind bislang weitgehend unbekannt.
Manche Robben wie See-Elefanten können durch diese Anpassungen bis zu zwei Stunden am Stück unter Wasser bleiben, ohne auftauchen zu müssen, und dabei Tiefen von bis zu 1500 Metern erreichen. Die meisten Arten erreichen solche Rekordwerte allerdings nicht. Dies hängt in erster Linie mit dem größeren Körpervolumen der See-Elefanten im Vergleich zu diesen Arten und der dadurch größeren Speicherkapazität für Sauerstoff zusammen. Die meisten Tauchgänge der Robben liegen innerhalb der aeroben Tauchgrenze der entsprechenden Art. Dadurch kann die Zeitdauer minimiert werden, für die sich eine Robbe nach jedem Tauchgang an der Wasseroberfläche erholen muss. Auf diese Weise können Robben bis zu 90 % ihres Tages unter Wasser verbringen.
Fortbewegung an Land
An Land wirken Robben eher unbeholfen. Hier bestehen Unterschiede zwischen den Ohrenrobben und Hundsrobben. Während erstere kräftige Hinterextremitäten behalten haben, mit denen sie den Körper stützen und auch Aktivitäten wie Körperpflege durchführen können, sind die Hinterbeine der Hundsrobben an Land weitgehend nutzlos, so dass sie auf dem Bauch „robben“ und sich vorwärts ziehen müssen.
Verbreitung und Lebensraum
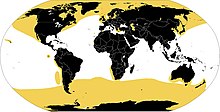

Nahezu alle Robben bewohnen das Meer. Da sie zu vielen Tätigkeiten immer wieder an Land kommen müssen, entfernen sie sich aber nicht allzu weit von der Küste und sind daher nicht auf hoher See anzutreffen.
Nur eine kleine Zahl von Robben tritt (1) fakultativ, (2) mit Präferenz als Unterart oder sogar (3) permanent als Endemit in Binnengewässern auf: (1) Der gemeine Seehund, überwiegend ein Meerestier, lebt in Kanada auch an Seeufern. (2) Zwei Unterarten der Ringelrobbe kommen hauptsächlich in Süßgewässern vor, die Ladogaringelrobbe (P. h. ladogensis) im russischen Ladogasee und die Saimaa-Ringelrobbe (P. h. saimensis) im finnischen Saimaa-See. (3) Zwei endemisch vorkommende Arten leben ausschließlich außerhalb des Meeres, die Baikalrobbe (P. sibirica) und die Kaspische Robbe (P. caspica). Die wahrscheinlichste Erklärung sind Binnenlandwanderungen der arktischen Ringelrobbe (Pusa hispida) während einer Kaltzeit etwa vor 400.000 Jahren, was ihrer genetischen Distanz zur Baikalrobbe entspricht. Dazu müssten die Vorfahren der Baikalrobbe 3800 km der Angara folgend von der Arktis zurückgelegt haben. Allerdings bestand während starker Kaltzeiten immer wieder eine Verbindung zum (derzeit nicht existierenden) Gletscherstausee, genannt Komisee, zuletzt vor etwa 60.000 bis vor 50.000 Jahren. Im Fall der Kaspischen Robbe besteht im jetzigen Interstadial keine Flussverbindung zwischen dem Kaspischen Meer und dem Arktischen Ozean, wohl gab es aber eine Verbindung zum Schwarzen Meer.
Der Großteil der Robben lebt in polaren und subpolaren Breiten. Die Meere der Arktis und Antarktis haben einen bemerkenswerten Arten- und Formenreichtum. Hier bilden viele Robben große Kolonien an den Küsten unbewohnter Inseln. Hingegen nimmt die Artenzahl zu den gemäßigten Meeren hin schnell ab, und in den Tropen gibt es fast keine Robben. Auch hier gibt es allerdings Ausnahmen, zum Beispiel die Mönchsrobben und den Galápagos-Seebär. Großregionen, in denen es überhaupt keine Robben gibt, sind die Küsten des tropischen Afrikas, die asiatischen Küsten des Indischen Ozeans und die Inselwelt des westlichen Pazifik. Kennzeichnend für fast alle Verbreitungsgebiete ist, dass die Wassertemperatur ganzjährig unterhalb von 20 °C bleibt; die Mönchsrobben bilden die einzige Ausnahme von dieser Regel.
An den Küsten von Nord- und Ostsee gibt es nur drei Robbenarten: den Seehund, die Kegelrobbe und die Ringelrobbe. Der Seehund ist in der Nordsee allgegenwärtig, in der Ostsee aber eine extreme Rarität; Kegelrobben-Kolonien findet man auf deutschem Boden auf dem Jungnamensand westlich von Amrum sowie auf Helgoland, außerhalb der Jungenaufzucht an allen Nordseeküsten sowie selten an vorpommerschen Ostseeküsten; die Ringelrobbe lebt im Finnischen und Bottnischen Meerbusen der Ostsee und gelangt nicht an deutsche Küsten.
Lebensweise

Im Gegensatz zu Walen und Seekühen, die vollkommen zum Wasserleben übergegangen sind, führen Robben eine amphibische Lebensweise. Die Paarung und die Jungenaufzucht findet an Land statt. Obwohl Robben auch an der Wasseroberfläche schlafen können, kommen sie oft zum Ruhen an die Küste.
Ernährung
Alle Robben sind Fleischfresser. Die Mehrzahl der Arten ernährt sich dabei von Fischen. Einige haben aber besondere Lebensweisen entwickelt: So bildet Krill die Nahrungsgrundlage des Krabbenfressers, das Walross sucht den Meeresgrund nach Schnecken und Muscheln ab, Tintenfische bilden einen Teil der Nahrung von See-Elefanten, während der Seeleopard Jagd auf Pinguine und kleinere Robben macht. Kleine Beutetiere werden meist bereits unter Wasser als Ganzes verschluckt, während größere Nahrung an die Wasseroberfläche gebracht wird, wo gegebenenfalls einzelne Stücke abgebissen werden. Besonders Jungtiere sind auf ein reichhaltiges Nahrungsangebot angewiesen, da sie aufgrund ihrer geringeren Körpergröße ein ungünstiges Verhältnis von Körperoberfläche zu -volumen haben und daher stärkere Wärmeverluste ausgleichen müssen. Gesunde ausgewachsene Tiere sind dagegen nicht zuletzt durch ihre unter der Haut gelegene Fettschicht in der Lage, auch längere Fastenzeiten zu überstehen.
Gastrolithen sind aus den Mägen von verschiedenen Robben- und Seelöwenarten bekannt. Die Funktion der Magensteine ist noch weitgehend unklar, plausibel sind sowohl die Zerkleinerung der Nahrung im Magen als auch die Verringerung des Auftriebs im Wasser.
Sozialverhalten
Die meisten Robben leben gesellig. Einzelgängerische Arten wie die Ross-Robbe sind die Ausnahme. Vor allem zur Jungenaufzucht finden sich Robben zu Kolonien zusammen, die in der Größe von einigen Individuen (Kegelrobbe) bis zu mehreren Millionen Tieren (Südliche Seebären) reichen können. Allerdings sind sehr große Kolonien infolge der massenhaften Abschlachtungen voriger Jahrhunderte selten geworden.
Lautverhalten
Robben kommunizieren lautlich mittels Klicklauten und singendem Pfeifen. Größere Robben entwickeln ein breites Spektrum von Lauten wie ein lang gezogenes tiefes Dröhnen. Von einer Robbenart (erstmals wahrgenommen bei dem Seehund Hoover, Boston Aquarium) ist bisher bekannt, dass sie die Laute des Menschen nachahmt.
Fortpflanzung
Die Fortpflanzungszeit liegt typischerweise im Frühling oder Frühsommer. Bei den meisten Arten etablieren die Männchen Territorien, von denen sie Konkurrenten fernzuhalten versuchen. Bei den dabei auftretenden Rivalenkämpfen haben schwächere Männchen unweigerlich das Nachsehen, so dass ein dominantes Männchen meist mehrere Partnerinnen für sich reklamieren kann.
Die meist vom Vorjahr trächtigen Weibchen treffen manchmal um Wochen später als die Männchen in der Kolonie ein. Sie suchen sich dort die besten Plätze für die Aufzucht ihres Nachwuchses aus, die in der Regel von den durchsetzungsstärksten Männchen besetzt sind; dort kommt ihr Junges zur Welt. Hundsrobben säugen es je nach Art für einige Tage oder mehrere Wochen und paaren sich dann nach der Entwöhnung mit dem Besitzer des Territoriums. Die Beziehung zwischen Mutter und Jungtier ist zu diesem Zeitpunkt praktisch beendet. Bei Ohrenrobben bleibt der Kontakt dagegen über die Paarung der Mutter hinaus bestehen, die meist etwa eine Woche nach der Geburt ihres Nachwuchs stattfindet. Dieser wird erst nach etwa vier bis sechs Monaten selbstständig, kann aber gelegentlich sogar noch nach der Geburt des nächsten Jungen mit Muttermilch versorgt werden.
Nach der Befruchtung entwickelt sich der Embryo nur bis zum Blastozysten-Stadium, einer noch undifferenzierten Hohlkugel aus Zellen. Erst nach einer Periode der Keimruhe nistet sich die Blastozyste in der mütterlichen Gebärmutterwand ein und bildet eine Plazenta aus. Nur durch dieses System können Robbenweibchen Geburt und erneute Befruchtung in demselben Zeitabschnitt bewerkstelligen.
Die Tragzeit beträgt bei Robben je nach Art acht bis fünfzehn Monate. Alle Arten bringen in der Regel ein Junges zur Welt, das entweder mit dem Kopf oder mit dem Schwanz voraus geboren wird. Zwillingsgeburten kommen vor, sind aber sehr selten; die Milch der Mutter reicht dann auch fast nie zur Aufzucht beider Tiere. Neugeborene besitzen ein spezielles flaumiges Geburtsfell, das Lanugo genannt wird und sich oft von der Fellfarbe der erwachsenen Tiere unterscheidet. Einige Hundsrobben-Arten wechseln es allerdings bereits in der Gebärmutter, während die meisten Ohrenrobben erst mit zwei bis drei Monaten ihr Jungenkleid ablegen. Alle Jungtiere haben noch keine ausreichende Fettschicht, um Auftrieb und Wasserabweisung zu gewährleisten. Obwohl sie theoretisch von Anfang an schwimmen könnten, gehen sie daher erst im Alter von einigen Wochen erstmals ins Wasser. Da die Jungtiere am Anfang ihres Lebens weitgehend schutzlos sind, geht das Wachstum sehr schnell vonstatten.
Fressfeinde und Lebenserwartung
Zu den Feinden der Robben gehören vor allem Haie und Schwertwale. Letztere haben sich in Patagonien so auf die Robbenjagd spezialisiert, dass sie sich sogar auf den Strand werfen, um die fliehenden Robben zu erbeuten. In der Arktis tritt der Eisbär als wichtiger Feind der Robben in Erscheinung, in der Antarktis der Seeleopard, der selbst eine Robbenart ist, die sich auf die Jagd auf ihre Verwandten spezialisiert hat.
Die meisten Arten haben eine Lebenserwartung von etwa dreißig Jahren. Das Walross wird oft älter als vierzig, viele Ohrenrobben nicht älter als zwanzig Jahre. Bei den in Herden lebenden Robben sterben die Männchen oft sehr viel früher als die Weibchen, weil sie sich bei den Revierkämpfen verausgaben oder schwere Verletzungen zuziehen. Auch die kleineren Robbenarten haben theoretisch eine hohe Lebenserwartung – die Seehunde zum Beispiel über 30 Jahre – die sie aber wegen natürlicher Feinde und menschlicher Einflüsse selten ausschöpfen können.
Stammesgeschichte

Moderne Arten
Traditionell wurden Robben entweder als eine eigenständige Ordnung angesehen oder sie wurden als Unterordnung der Wasserraubtiere den Landraubtieren gegenübergestellt. Zumindest die letztere Einordnung ist heute unüblich. Die Robben haben sich aus Landraubtieren entwickelt, genauer gesagt aus Hundeartigen; sie sind daher auch innerhalb der Hundeartigen anzusiedeln.
Aufgrund morphologischer Untersuchungen gingen manche Zoologen noch in den 1990ern davon aus, dass die Robben zwei verschiedene Entwicklungslinien bilden. So meinte man, dass die Ohrenrobben von bärenähnlichen Ahnen und die Hundsrobben von otterartigen Vorfahren abstammten. Demnach hätten sich die beiden Robbengruppen unabhängig voneinander entwickelt. Dieser Hypothese folgend wären Robben polyphyletisch, also ein reines Formtaxon, das keine Berechtigung als systematische Gruppe der Säugetiere hätte.
In molekulargenetischen Analysen wurde allerdings seit den 1990ern diese Hypothese zurückgedrängt. Olaf R. P. Bininda-Emonds und A.P. Russell führten 1996 starke Belege für eine Monophylie der Robben an; spätere Studien haben ihre Ergebnisse bestätigt.
Seit dem 19. Jahrhundert wird eine Verwandtschaft der Robben mit den Bären und ihren Verwandten für wahrscheinlich gehalten. Allerdings ist die genaue Position der Robben im zoologischen System noch unklar, so dass auch die Schwestergruppe der Robben noch nicht zweifelsfrei bekannt ist.
Bei Malcolm C. McKenna und Susan K. Bell tauchen die Robben als Schwestergruppe der Bären auf und dies innerhalb eines übergeordneten Taxons Ursida:
| Ursida |
|
||||||||||||
Die Amphicyonidae sind eine ausgestorbene Gruppe, die vom Eozän bis zum Miozän lebte; die Ursoidea umfassen die Bären und die ebenfalls ausgestorbenen Hemicyonidae (Eozän bis Pliozän); und Phocoidea ist der von Malcolm C. McKenna und Susan K. Bell synonym für Pinnipedia verwendete Begriff. Die hier gezeigte Hypothese ist am weitesten verbreitet, doch gibt es auch widersprechende Ansichten, in denen beispielsweise die Marder oder der Kleine Panda als Schwestergruppen der Robben angesehen werden.
Fossile Vertreter

Die ersten fossil erhaltenen robbenartige Tiere stammen aus dem späten Oligozän vor etwa 27 bis 25 Millionen Jahren, darunter der otterähnliche Puijila und die Gattungen Enaliarctos und Pacificotaria, die in die Familie Enaliarctidae gestellt werden. Diese lange Zeit rätselhafte Gruppe wurde in jüngerer Zeit durch Funde vollständiger Skelette besser bekannt. Die Tiere der Gattungen Enaliarctos und Pacificotaria waren bereits robbenähnlich, hatten aber noch zum Laufen an Land geeignete Vorder- und Hinterbeine. Ob die Enaliarctidae oder die ebenfalls ausgestorbenen Tiere der Gattung Pteronarctos bereits echte Robben waren, ist eine Frage der Definition. A. Berta und A. R. Wyss verneinten dies 1994 und fassten Robben und Enaliarctidae zu einem höheren Taxon Pinnipedimorpha zusammen:
| Pinnipedimorpha |
|
||||||||||||||||||||||||
Der biogeografische Ursprung der Robben lag sehr wahrscheinlich im Nordpazifik, vermutlich an der Westküste des nordamerikanischen Kontinents. Aus diesem Gebiet stammen vermutlich auch die modernen Taxa der Ohren- wie Hundsrobben, die erstmals im Miozän auftraten. Durch Fossilien ist dies allerdings nur für die Ohrenrobben und Walrosse belegt, deren früheste Arten allesamt aus nordpazifischen Fundstätten stammen; die ersten Hundsrobben-Fossilien sind dagegen erst aus ehemals atlantischen Gewässern vor der Ostküste der USA bekannt.
Systematik

Nach dem Aussterben des Japanischen Seelöwen und der Karibischen Mönchsrobbe existieren noch 34 rezente Robbenarten. Es werden traditionell drei Familien unterschieden:
- Hundsrobben (Phocidae) besitzen keine äußerlich sichtbaren Ohren; ihre Beinflossen sind nach hinten ausgerichtet, so dass sie bei der Fortbewegung an Land nicht zum Einsatz kommen können. Im Wasser liefern sie im Wechselschlag dagegen den Vortrieb der Tiere. Nach dem Aussterben der Karibischen Mönchsrobbe existieren noch 18 Arten. Die Hundsrobben umfassen eine Vielzahl kleinerer Robben wie etwa Seehunde oder Kegelrobben, aber auch die riesigen See-Elefanten.
- Ohrenrobben (Otariidae) haben kleine äußerliche Ohren und können ihre Beinflossen nach vorne unter den Körper drehen, um sich so besser an Land bewegen zu können. Bei ihnen sind im Wasser die Vorderflossen die Hauptquelle des Vortriebs. Die 15 Arten der Gruppe sind vor allem große koloniebildende Robben wie Seelöwen oder Seebären.
- Walrosse (Odobenidae), heute nur noch durch eine Art vertreten, zeichnen sich in erster Linie durch die auffälligen Stoßzähne, die stark vergrößerten Eckzähne des Oberkiefers, aus. Sie können ihre Hinterflossen, die eher jenen der Hundsrobben ähneln, wie Ohrenrobben unter den Körper stellen.
Die Ohrenrobben werden wegen ihrer sichtbaren äußeren Ohren gelegentlich als die „primitivere“ Gruppe angesehen, doch die fossilen Belege der Hundsrobben sind ebenso alt wie die der Ohrenrobben.
Ohrenrobben und Walrosse werden meistens als eng verwandte Kladen angesehen, obwohl die Walrosse in zahlreichen Merkmalen zwischen beiden Gruppen vermitteln. Malcolm C. McKenna und Susan K. Bell haben in Widerspruch zu dieser weit verbreiteten Ansicht die Walrosse als Unterfamilie der Hundsrobben eingestuft. Dies ist jedoch eine Minderheitensicht, denn die meisten Zoologen sehen starke Belege für ein gemeinsames Taxon von Ohrenrobben und Walrossen, manchmal Otarioidea genannt:
| Robben (Pinnipedia) |
|
||||||||||||
Walrosse (Odobenidae)
|
Menschen und Robben

Die Beziehung zwischen Menschen und Robben ist stark von der Region abhängig, in der sich eine menschliche Kultur befand. Sie begann sehr wahrscheinlich schon in der Altsteinzeit, aus der Gravuren in Robbenknochen und -zähnen bekannt sind. Aus der Kultur der Eskimos sind Robben bis heute kaum wegzudenken, bilden sie doch nicht nur eine bedeutende Nahrungsquelle, sondern liefern auch Haut für Leder und das Robbenfell für Pelze. Früher wurden auch Knochen zur Werkzeugherstellung, Sehnen für Bogen und Fett für wärme- und lichtspendende Öllampen genutzt.
Auch anderen Völkern dienten Robben als Nahrungsgrundlage. So erlegten die Māori der Südinsel Neuseelands und die Aborigines australischer Südküsten Seebären und nutzten sie in ähnlich vielfältiger Weise. Seebären waren auch eine beliebte Beute der Ureinwohner des südamerikanischen Feuerland.
Dagegen hatten die antiken Reiche des Mittelmeerraums wenig Berührung mit Robben, da hier nur eine Art, die Mittelmeer-Mönchsrobbe, beheimatet ist, die wahrscheinlich schon im Altertum eine Seltenheit war. Aristoteles lieferte eine Beschreibung dieser Robbe, die somit der früheste in schriftlicher Überlieferung erhaltene Vertreter der Tiergruppe überhaupt ist. Dass an Küsten liegende Mönchsrobben die Vorlage für die Sirenen der griechischen Mythologie waren, ist denkbar, bewegt sich aber im Bereich der Spekulation.
Keine der angesprochenen Subsistenz-Jagden hatte vermutlich einen wesentlichen langfristigen Einfluss auf die Bestände der betroffenen Robbenpopulationen. Robben können an Plastiktüten sterben, weil diese für Quallen gehalten werden.
Moderne Massenjagd

Die große Zeit der Robbenjäger begann im späten 18. Jahrhundert, obwohl schon seit dem 16. Jahrhundert Schiffe einzig zum Zweck des Schlachtens von Robben ausgesandt wurden. Im gesamten 19. Jahrhundert waren im Nord- und Südpolarmeer Schiffe unterwegs, die an Inseln anlegten, um die dort in Kolonien lebenden Großrobben zu erschlagen. Besonders betroffen waren dabei die Seebären, deren Fell man als besonders wertvoll erachtete. Die Robbenjagden erreichten ein Ausmaß, das unter den Ausrottungen im Tierreich seinesgleichen sucht. So lagerten an den Juan-Fernández-Inseln in jedem Sommer etwa fünfzehn Schiffe gleichzeitig, deren Besatzungen jährlich 250.000 an den Küsten lagernde Seebären erschlugen und somit eine der größten Tierkolonien der Welt binnen fünfzehn Jahren restlos vernichteten. Der Antarktische Seebär, der zu etlichen Millionen Tieren an den Küsten subantarktischer Inseln beheimatet war, wurde von kreuz und quer durch das Südpolarmeer fahrenden Robbenjägern zwischen 1800 und 1830 beinahe ausgerottet. Die Südlichen Shetlandinseln wurden beispielsweise erst 1819 entdeckt, doch schon zwei Jahre später waren die fünfhunderttausend hier lebenden Seebären bis auf das letzte Exemplar getötet. Dass bei diesen Ausrottungsexzessen nur eine Art, die Karibische Mönchsrobbe (Monachus tropicalis), ausgestorben ist, ist eine erstaunliche Tatsache. Allerdings sind fast alle Robben in ihrem Bestand stark zurückgegangen, und manche Arten, die früher gewaltige Kolonien bildeten, leben heute nur noch in winzigen Grüppchen an den Küsten.
Heute werden meistens andere Robbenarten getötet, denn die letzten verbliebenen Seebären stehen größtenteils unter Schutz. Es ist vor allem die Sattelrobbe, deren Jungtiere zur Gewinnung des Fells getötet werden. Diese Tötungen werden alljährlich von Protesten von Tierschützern begleitet, die öffentlichkeitswirksam mit den Gesichtern von Robbenbabys werben. Die traditionelle Jagd erwachsener Tiere durch die Eskimos wird im Gegensatz dazu meist auch von Naturschützern akzeptiert. Die Befürworter rechtfertigen die Notwendigkeit zur Dezimierung der Robben unter anderem mit dem Argument, dass eine nicht kontrollierte Robbenpopulation die Fischbestände plündere; Meeresbiologen gehen allerdings davon aus, dass der Rückgang der Fischschwärme nicht von den wenigen verbliebenen Robben, sondern von den Hochseeflotten der Industriestaaten verursacht wird.
1972 wurde als Zusatzprotokoll zum Antarktisvertrag das Übereinkommen zur Erhaltung der Antarktischen Robben (Convention on the Conservation of Antarctic Seals), kurz (CCAS) geschlossen.
Am 5. Mai 2009 verabschiedete das Europaparlament ein Handelsverbot für zahlreiche aus Robben hergestellte Produkte wie Robbenfelle sowie unter Verwendung von Robbenöl hergestellte Omega-3-Kapseln. Eng begrenzte Ausnahmen gestatten einen Handel mit durch Eskimos erlegten Robben, für die ein Herkunftsnachweis erforderlich wird. Das Verbot sollte vor der nächsten Jagdsaison 2010 in Kraft gesetzt werden. Ein Hauptargument ist dabei nicht die wirtschaftliche Nutzung der Robbenbestände an sich, sondern die als nicht tierschutzgerecht eingestufte Art und Weise, in der die Tiere erschlagen werden. Die hauptsächlich am Handel mit diesen Produkten beteiligten Staaten Kanada und Norwegen legten bei der Welthandelsorganisation Beschwerde dagegen ein und erreichten damit, dass das Embargo mindestens bis zum 7. September 2010 ausgesetzt war.
In der EU gilt unmittelbar die Verordnung (EG) Nr. 1007/2009 des Europäischen Parlaments und des Rates vom 16. September 2009 mit einem Verbot des Inverkehrbringens für Robben (und Teile von ihnen wie Felle) und für Erzeugnisse aus Robben (wie Öle). Das Inverkehrbringen ist hier definiert als Einfuhr in den gemeinsamen Markt, durch die ein entgeltliches Bereitstellen für Dritte erfolgt; es sind einige Ausnahmen vorgesehen, die an die Art der Robbenjagd und ihren Nachweis anknüpfen. Die Durchführung des Verbots regelt in Deutschland das Tiererzeugnisse-Handels-Verbotsgesetz.
Zoos und Zirkusse
Auch in Zoos und gelegentlich noch in Zirkussen werden Robben gehalten. Oft sind im Zoo die Fütterungen der Robben besondere Zuschauermagneten. Berühmt geworden ist das Walross Antje aus dem Tierpark Hagenbeck, das zum Logo des NDR wurde. In Zirkussen, Delfinarien und anderen Tiershows wird bevorzugt der Kalifornische Seelöwe gezeigt, der sich zu Kunststücken mit Bällen und Reifen dressieren lässt.
Viele Tierschützer bemängeln allerdings, dass sich Robben im Zoo nicht artgerecht halten lassen – dies gilt vor allem für die häufig in Zoos gehaltenen Ohrenrobben, die in freier Wildbahn in großen Kolonien leben und bei denen Revierkämpfe zum Verhaltensrepertoire gehören. Insbesondere die Wasserbecken, in denen die Tiere gehalten werden, gelten als zu flach, um den Tieren annähernd natürliche Gegebenheiten zu bieten; daneben ist das Wasser oft gechlort.
Militärische Nutzung
Relativ neu sind die Bemühungen der US-amerikanischen Marine, dressierte Seelöwen in die militärische Planung einzubeziehen. Die Tiere werden darauf trainiert, Seeminen aufzuspüren, vor feindlichen Tauchern zu warnen und in tiefem Wasser verloren gegangene Objekte wiederzubeschaffen. Ähnliche Experimente führte die schwedische Marine im Zweiten Weltkrieg durch.
Literatur
- Ronald M. Nowak: Walker’s mammals of the world. 6. Auflage. Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9 (englisch).
- Bernhard Grzimek: Grzimeks Tierleben. Band 12 (Säugetiere 3). Bechtermünz, Augsburg 1979, 2000, ISBN 3-8289-1603-1.
- C. McKenna, Susan K. Bell: Classification of Mammals. Above the Species Level. Columbia University Press, New York 2000, ISBN 0-231-11013-8.
- Rüdiger Wandrey: Die Wale und Robben der Welt. Franckh-Kosmos, Stuttgart 1997, ISBN 3-440-07047-6.
- A. Berta, A. R. Wyss: Pinniped phylogeny. In: Contributions in marine mammal paleontology honoring Franck C. Whitmore, Jr. Proceedings of the San Diego Society of Natural History. 29, 1994, S. 33–56. ISSN 1059-8707
- Olaf R. P. Bininda-Emonds, A. P. Russell: A morphological perspective on the phylogenetic relationships of the extant phocid seals (Mammalia: Carnivora: Phocidae). In: Bonner zoologische Monographien. 41, 1996, S. 1–256. ISSN 0302-671X