
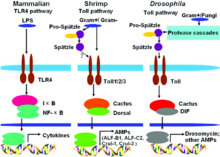
Toll-like-Rezeptoren (kurz TLR, englisch toll-like receptor) sind Strukturen des angeborenen Abwehrsystems (innate immunity) und gehören zu einer Gruppe von Rezeptoren, den PRRs (Pattern-Recognition Receptors). Sie dienen der Erkennung von PAMPs (Pathogen-Associated Molecular Patterns). Das sind Strukturen, welche ausschließlich auf oder in Krankheitserregern vorkommen, und steuern entsprechende Aktivierungen von Genen. Hierdurch wird die Aktivierung des „antigen-spezifischen erworbenen Immunsystems“ (antigen-specific acquired immunity) eingeleitet und moduliert. Durch die „toll-like receptors“ vermag das angeborene Abwehrsystem zwischen „selbst“ und „nicht selbst“ zu unterscheiden. TLRs werden im Allgemeinen in dendritischen Zellen und Makrophagen exprimiert, wenig bis gar nicht in Epithelzellen.
Der Name Toll-like-Rezeptor (in der deutschsprachigen Literatur als Signaltransduktions-vermittelnde PRRs oder selten auch als Toll-artiger Rezeptor bezeichnet) ist von einem Protein bei Drosophila melanogaster abgeleitet, über dessen Entdeckung die Forschungsgruppe um die Nobelpreisträgerin Christiane Nüsslein-Volhard derart begeistert war, dass sie es humorvoll nach dem deutschen Ausdruck toll nannte. Der Moment, der dem Toll-Gen der Fruchtfliege seinen Namen gab, war, als sie ihrem Kollegen Eric Wieschaus gegenüber an einem Doppelmikroskop saß, das zwei Personen gleichzeitig die Untersuchung desselben Objekts erlaubt. „Als wir eines Tages eine Embryonenmutante sahen, deren Entwicklung ventralisiert war, waren wir beide vollkommen überrascht und haben spontan ‚toll‘ gerufen. Bis dahin kannten wir nur dorsalisierte Embryonen“. TLRs bestehen aus Proteinen, die Toll ähneln, also toll-like sind.
Seit der Entdeckung des ersten Toll-like-Rezeptoren Mitte der 1990er Jahre sind immer wieder neue Varianten in Menschen und Tieren entdeckt worden. TLRs finden sich in allen Vertebraten, aber auch in einfacheren Organismen wie zum Beispiel in Drosophila melanogaster, was nahelegt, dass es sich um ein evolutionär sehr altes System handelt. Die meisten Spezies verfügen über mehr als zehn verschiedene (bekannte) TLRs, wobei manche Typen z. B. in der Maus, jedoch nicht im Menschen vorkommen.
TLRs erkennen verschiedene funktionale Bestandteile von Viren, Bakterien und Pilzen und können so biochemische Reaktionsketten in den Zellen auslösen, die der Abwehr dieser Krankheitserreger dienen.
Entdeckung der TLRs

Als Mikroganismen erstmals als die Ursache für Infektionskrankheiten identifiziert wurden, war klar, dass mehrzellige Organismen diese erkennen können müssen und dass es dafür notwendig ist, für Mikroorganismen typische Molekülstrukturen zu erkennen. Eine große Menge an Literatur, die den Großteil des 20. Jahrhunderts umfasste, widmet sich den Schlüsselmolekülen und ihren Rezeptoren. Vor mehr als 100 Jahren prägte Richard Pfeiffer, ein Schüler von Robert Koch, den Begriff Endotoxin, um eine Substanz, die von gramnegativen Bakterien produziert wurde und bei Tierversuchen zu Fieber und Schockzuständen führte, zu benennen. In den folgenden Jahrzehnten wurden die Endotoxine chemisch charakterisiert und als Lipopolysaccharide (LPS), die von den meisten gramnegativen Bakterien produziert werden, identifiziert. Es wurde gezeigt, dass auch andere Moleküle (bakterielle Lipopeptide, Flagelline und nicht methylierte DNA) zu einer Immunantwort führen. Logischerweise wurde daraus geschlossen, dass es Rezeptoren geben muss, die in der Lage sind, eine Immunantwort für solche Molekülstrukturen herbeizuführen. Diese wurden jedoch viele Jahre lang nicht gefunden.
In der Mitte der 1990er wurde durch Forschungen im Bereich der Entwicklungsbiologie von Drosophila melanogaster eher zufällig erkannt, dass Toll-negative Mutanten sehr anfällig gegen Pilzbefall sind. Diese Beobachtung leitete eine gezielte Suche nach ähnlichen Proteinen in Säugerzellen ein. 1994 konnte von Nomura und Kollegen der erste menschliche TLR gefunden werden, der 1996 von Taguchi und Kollegen einem Chromosom zugeordnet werden konnte. Dies zeigt, dass es sich bei der über Toll-like Receptor vermittelten Immunantwort um eine evolutionär sehr alte Form handelt, die genetisch hochkonserviert ist. Da die Rolle der TLRs bei der Immunabwehr zum damaligen Zeitpunkt noch nicht bekannt war, wurde angenommen, dass TLR-1 in der Entwicklungsbiologie der Säugetiere eine Rolle spielen würde. 1997 zeigten Charles Janeway und Ruslan Medzhitov, dass ein Toll-like Receptor, wenn er künstlich an entsprechende Antikörper gebunden wird, bestimmte Gene aktivieren kann, die für eine adaptive Immunantwort nötig sind. Die Funktion des TLR-4 als LPS-Rezeptor wurde von Bruce A. Beutler und Kollegen entdeckt. Im Laufe der Zeit wurden auch die Liganden der anderen TLRs bestimmt. Shizuo Akira nahm dabei eine zentrale Rolle ein.
Struktur und Liganden
Die gemeinsamen Strukturmerkmale aller Toll-like-Rezeptoren sind die N-terminalen leucinreichen LRR-Sequenzen (Leucine-rich Repeats) und die TIR-Domain (Toll/IL-1R homology domain). Die unterschiedlichen TLRs können jeweils unterschiedliche PAMPs (Pathogen Associated Molecular Patterns) durch direkte Interaktion mit der jeweiligen Membranoberfläche des Krankheitserregers identifizieren. TLR-3, TLR-7, TLR-8 und TLR-9 befinden sich an der Membran von Endosomen bzw. Endolysosomen, während die restlichen Toll-like-Rezeptoren an der Zellmembran sitzen.
TLR-2 erkennt viele Komponenten von Bakterien, Mykoplasmen, Pilzen und Viren. Hierzu gehören auch die Lipoproteine der Bakterien und Mykoplasmen. Die Erkennung erfolgt, indem TLR2 entweder mit TLR1 oder TLR6 eine Heterodimer bildet. Die entstehenden TLR-1/TLR-2- und TLR-6/TLR-2-Komplexe erkennen dabei Triacyl- bzw. Diacyllipoproteine. Die Aktivierung von TLR2 führt zur Ausschüttung einer Reihe von Zytokinen. Alpha-Interferon (IFN-α) wird dabei nur teilweise freigesetzt. Allgemein wird vermutet, dass die Ausschüttung von IFN-α vom Zelltyp abhängig ist. TLR-10 ähnelt in seiner Primärstruktur (Sequenzübereinstimmung) TLR-1 und TLR-6, der Ligand ist jedoch noch nicht bekannt. TLR-4 kann Lipopolysaccharid (LPS) auf der Zelloberfläche zusammen mit MD2 (myeloid differentiation factor 2) erkennen. LPS ist ein Stoff aus der äußeren Membran von gramnegativen Bakterien und wird in Tiermodellen zur eingeschränkten Simulation von akuten Infektionen verwendet. Bei der Detektion von LPS arbeiten zwei TLR4-MD2-LPS-Komplexe zusammen und formen ein TLR4-Homodimer.
TLR5 wird hauptsächlich in der Lamina propria exprimiert, wo es bakterielles Flagellin erkennt. Als Immunantwort darauf induziert TLR-5 die Ausdifferenzierung von B-Zellen in lgA-produzierende Blutzellen und von T-Zellen in antigenspezifische Th17- und Th1-Zellen. TLR-11 das nur in der Maus, aber nicht im Menschen vorkommt, zeigt große Ähnlichkeiten zu TLR-5. Es erkennt ein Profilin-ähnliches Molekül, das vom intrazellulären Protozoon Toxoplasma gondii abstammt. Eine Reihe von TLRs, darunter TLR-3, TLR-7, TLR-8 und TLR-9 erkennen aus Viren oder Bakterien stammende Nukleinsäuren. Die Aktivierung dieser TLRs führt zur Bildung von Alpha-Interferon und anderen entzündungsauslösenden Zytokinen. TLR-3 entdeckt virale doppelsträngige RNA im Endolysosom. Hierbei bindet die dsRNA an das N-terminale und das C-terminale Ende der LRR-Sequenz.
| Rezeptor | Liganden | Ursprung der Liganden | Adapterprotein des TLR | Zelluläre Lokalisation | Zelltypen |
|---|---|---|---|---|---|
| TLR-1 | Verschiedene Triacyl-Lipopeptide | Bakterielles Lipoprotein | MyD88/MAL | Zelloberfläche |
|
| TLR-2 | Verschiedene Glycolipide | Bakterielle Peptidoglykane | MyD88/MAL | Zelloberfläche |
|
| Verschiedene Lipopeptide und Proteolipide | Bakterielle Peptidoglycane | ||||
| Lipoteichonsäure | Gram-positive Bakterien | ||||
| HSP70 | Wirtszellen | ||||
| Zymosan (Beta-Glucan) | Pilze | ||||
| Zahlreiche Andere | |||||
| TLR-3 | Doppelsträngige RNA, Poly I:C | Viren | TRIF | Zellkompartiment |
|
| TLR-4 | Lipopolysaccharide, Eritoran | Gram-negative Bakterien | MyD88/MAL/TRIF/TRAM | Zelloberfläche |
|
| Mehrere Hitzeschockproteine | Bakterien und Wirtszellen | ||||
| Fibrinogen | Wirtszellen | ||||
| Heparansulfatfragmente | Wirtszellen | ||||
| Hyaluronsäurefragmente | Wirtszellen | ||||
| Nickel | |||||
| Verschiedene Opioide | |||||
| TLR-5 | Bakterielles Flagellin | Bakterien | MyD88 | Zelloberfläche |
|
| Profilin | Toxoplasma gondii | ||||
| TLR-6 | Verschiedene Diacyl-Lipopeptide | Mycoplasma | MyD88/MAL | Zelloberfläche |
|
| TLR-7 | Imidazochinolin | Niedermolekulare synthetische Stoffe | MyD88 | Zellkompartiment |
|
| Loxoribin (ein Guanosinanalogon) | |||||
| Bropirimin | |||||
| Imiquimod, Resiquimod | |||||
| Einzelsträngige RNA | RNA-Viren | ||||
| TLR-8 | Niedermolekulare synthetische Stoffe, RNase-prozessierte virale und bakterielle RNA, Resiquimod | MyD88 | Zellkompartiment |
|
|
| TLR-9 | Unmethylierte CpG-Oligonukleotide (DNA) | Bakterien, DNA-Viren | MyD88 | Zellkompartiment |
|
| TLR-10 | Triacylierte Lipopeptide | unbekannt | Zelloberfläche |
|
|
| TLR-11 | Profilin | Toxoplasma gondii | MyD88 | Zellkompartiment |
|
| TLR-12 | Profilin | Toxoplasma gondii | MyD88 | Zellkompartiment |
|
| TLR-13 | Bakterielle ribosomale RNA-Sequenz CGGAAAGACC (unmethyliert) | Viren, Bakterien | MyD88, TAK-1 | Zellkompartiment |
|
Die intrazelluläre Signalkaskade

Die Chemoattraktorproteine „C3a“ und „C5a“ des Komplementsystems entstehen durch proteolytische Spaltung aus den inaktiven Vorstufen C3, bzw. C5. Aktiviert locken sie Makrophagen und neutrophile Granulozyten an. Diese Phagozyten haben auf ihrer Oberfläche Rezeptoren vom TLR-Typ. Die TLRs reagieren auf bakterielle Proteoglykane bzw. Lipopolysaccharide (LPS), DNA und RNA. Sie lösen in ihren Trägerzellen eine Signalkaskade aus, die schließlich zur Stimulation der Infektionsabwehr führt. Nach Erkennen dieser bakteriellen Oberflächenstrukturen auf extrazellulärer Seite, werden intrazellulär – durch TLR – Signalkaskaden ausgelöst.
Die Erkennung von PAMPs (Pathogen Associated Molecular Patterns) durch TLRs führt zu einer Erhöhung der Transkriptionsrate bestimmter Gene, je nachdem welche TLRs und welche Zelltypen beteiligt sind. Der Unterschied zwischen den von den einzelnen TLRs aktivierten Signalkaskaden kann zumindest teilweise durch die TIR-domain enthaltenden Adaptormoleküle (TIR domain-containing adaptor molecules) erklärt werden. Es gibt dabei fünf TIR domain-containing adaptor molecules, darunter MyD88, TRIF/TICAM-1 (TIR-domain-containing adaptor inducing IFN-β), TIRAP/Mal, TRAM (TRIF-related adaptor molecule), und SARM (Sterile-alpha and Armadillo motif-containing protein). TLR-Signalketten werden abhängig von der Beteiligung der Adaptormoleküle MyD88 und TRIF grob in zwei unterschiedliche Reaktionsketten unterteilt.
Dabei kommt es zur Phosphorylierung und somit Aktivierung intrazellulärer Kinasen, deren Aufgabe in der Phosphorylierung intrazellulärer Inhibitoren von Transkriptionsfaktoren besteht. Als Erstes bindet das Adapterprotein MyD88 an den zytoplasmatischen Abschnitt des TLR. Als Folge bindet nun die IL-1-Rezeptor-assoziierte Kinase (IRAK) an MyD88 und aktiviert sich durch Autophosphorylation selbst. Über weitere Einzelschritte kommt es schließlich zur Aktivierung des Transkriptionsfaktors NF-κB, der daraufhin in den Zellkern transloziert und dort die Expression von Genen für TNFα, IL-1, IL-12 und E-Selektin reguliert.
Literatur
- K. Takeda, S. Akira: Toll-like receptors in innate immunity. In: International immunology, Band 17, Nummer 1, Januar 2005, ISSN 0953-8178, S. 1–14, doi:10.1093/intimm/dxh186, PMID 15585605 (Review).
- Luke A. J. O’Neill: Das immunologische Frühwarnsystem. Spektrum der Wissenschaft, August 2005, S. 68–75
- S. E. Turvey, T. R. Hawn: Towards subtlety: understanding the role of Toll-like receptor signaling in susceptibility to human infections. In: Clinical immunology (Orlando FL), Band 120, Nr. 1, Juli 2006, ISSN 1521-6616, S. 1–9, doi:10.1016/j.clim.2006.02.003, PMID 16563867 (Review).
- Nicola Siegmund-Schultze: Deutsches Ärzteblatt: Toll-like-Rezeptoren: Neue Zielstruktur für immunstimulierende Medikamente. In: Deutsches Ärzteblatt. Band 104, Nr. 16. Deutscher Ärzte-Verlag, 20. April 2007, S. A-1072 / B-954 / C-908 (Online).